promoter
「promoter」の意味・「promoter」とは
「promoter」は英語の単語で、日本語では「プロモーター」と訳される。主に二つの意味を持つ。一つ目は、何かを推進する人、または団体を指す。例えば、新製品の宣伝活動を行うマーケティング担当者や、特定のイベントを企画・運営する人物などがこれに該当する。二つ目は、遺伝子の機能を制御するDNAの一部を指す。この場合、プロモーターは遺伝子の転写を開始する場所で、転写因子が結合する部位である。「promoter」の発音・読み方
「promoter」の発音は、IPA表記では/prəˈmoʊtər/となる。IPAのカタカナ読みでは「プラモウター」となる。日本人が発音するカタカナ英語では「プロモーター」と読む。この単語は発音によって意味や品詞が変わる単語ではない。「promoter」の定義を英語で解説
A promoter is a person or organization that helps plan and conduct certain events, campaigns, or activities. In genetics, a promoter is a region of DNA that initiates transcription of a particular gene. It's where transcription factors bind to start the transcription process.「promoter」の類語
「promoter」の類語としては、「advocate」、「sponsor」、「backer」などがある。これらの単語も同様に、何かを推進する人や団体を指す言葉である。しかし、それぞれ微妙なニュアンスの違いがある。例えば、「advocate」は主に意見や考えを積極的に支持・推進する人を指し、「sponsor」は金銭的な支援を行う人や団体を指す。「promoter」に関連する用語・表現
「promoter」に関連する用語としては、「promotion」、「promote」、「promotional」などがある。「promotion」は「昇進」や「宣伝」、「promote」は「昇進させる」や「宣伝する」という意味を持ち、「promotional」は「宣伝の」を意味する形容詞である。「promoter」の例文
1. The promoter is responsible for organizing the concert.(プロモーターはコンサートの組織を担当している。)2. He is a promoter of environmental conservation.(彼は環境保全の推進者である。)
3. The promoter region controls the transcription of the gene.(プロモーター領域は遺伝子の転写を制御する。)
4. She works as a promoter for a new product.(彼女は新製品のプロモーターとして働いている。)
5. The promoter failed to attract enough audience to the event.(プロモーターはイベントに十分な観客を引きつけることができなかった。)
6. The promoter was arrested for fraud.(プロモーターは詐欺で逮捕された。)
7. The promoter region is a crucial part of gene regulation.(プロモーター領域は遺伝子調節の重要な部分である。)
8. The promoter has a reputation for being reliable.(そのプロモーターは信頼性があると評判である。)
9. The promoter is planning a series of promotional events.(プロモーターは一連の宣伝イベントを計画している。)
10. The promoter was criticized for the poor organization of the event.(プロモーターはイベントの組織が不十分だったと批判された。)
プロモーター【promoter】
プロモーター
| 遺伝子名DNARNAの配列や構造など: | プライマーRNA プロセス型偽遺伝子 プロトがん遺伝子 プロモーター ホメオティツク遺伝子 ホメオボックス遺伝子 ホルモン応答配列 |
プロモーター
プロモーター
出典: フリー百科事典『ウィキペディア(Wikipedia)』 (2022/11/24 03:16 UTC 版)

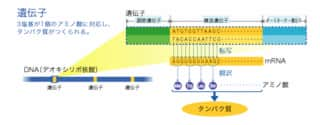
プロモーター(Promoter)とは転写(DNA からRNA を合成する段階)の開始に関与する遺伝子の上流領域[注釈 1]を指す。プロモーターに基本転写因子が結合して転写が始まる。
原核細菌のプロモーター
主に大腸菌の研究からプロモーターの中でも特に保存されている部分的配列(配列要素、element)が明らかになった[注釈 2]。これらの配列要素はRNAポリメラーゼが実際に結合する領域(大腸菌の場合60 bp)にはない[1]。
転写開始点
転写開始点は90%以上の場合でプリン塩基である[1]。CATという塩基配列の中央であることが比較的多いが、よく保存されているとは言い難い。
-35ボックスと-10ボックス
真正細菌のプロモーターにはRNAポリメラーゼを強力に引き寄せる2つの仕組みがある。一つは転写開始点から35 bp上流にある-35ボックス (-35領域または-35配列 とも)、もう一つは10 bp、より正確には-18~-9上流[1]の-10ボックス(-10領域または-10配列とも。当初は発見者のデビット・プリブノーにちなんでプリブノーボックスとも呼ばれていた[2])である[注釈 1]。2つは15~19 nt[3]、例外的に長いもので20 nt離れる[1]。
現在、数千ものプロモーターが調べられ、これら各ボックスにおける典型(共通配列、コンセンサス配列)が明らかになっている。共通配列とは現れる頻度の顕著な塩基配列であり、下図1に示す。図2に、各構成塩基ごとの存在する確率を記す。各ボックスが共通配列に完全に一致すると転写開始は劇的に頻発するだろう。実際には一致する例は少ないが、類似度合いの大きさがプロモーターの強さ(転写の頻度)となる。実際、各ボックスを典型に近づける突然変異は優勢変異といい、プロモーターを強力にして転写開始を促す[2]。逆に、類似性を下げる変異である劣勢変異を受けたプロモーターは弱い[2]。劣性変異の様相は受けるボックスによって変わり、-35ボックスの場合は閉鎖型複合体を形成する速度を減少させる[4]。-10ボックスでは、閉鎖型複合体に問題はないがそこから開放型複合体になるのは遅い。このことから-35ボックスはRNAポリメラーゼに認識される部位、-10ボックスは二重らせんをほどくのに重要な部位だと考えられている。
例外的に、一方または両方のボックスを持たないプロモーターも存在する[4]。この場合、補助因子がRNAポリメラーゼの認識を助ける。また、両ボックス以外の、最初に転写される+1~+30の領域もRNAポリメラーゼとDNAの結合の安定性に影響する[4]。
大腸菌の中には、-35ボックスの代わりにいわゆる「延長した-10ボックス」を持つものもいる[5]。この配列とRNAポリメラーゼとの間の接触が-35ボックスの欠如を補う。例として、ガラクトース代謝に関与するgal 遺伝子群がある。
このほか、-10ボックスのすぐ下流にRNAポリメラーゼと結合する弁別要素が発見された。酵素とプロモーターの作る複合体[注釈 3]の安定性に影響を与える。プロモーターの各要素は サブユニットの一つであるσ因子と結合してRNAポリメラーゼを誘導する。どのように結合するのかはRNAポリメラーゼ#σ因子に詳述している。
上流 プロモーター mRNAに転写される領域 下流 5'----------|TTGACa(-35)|---------|TAtAaT(-10)|----------------------|T|------------3' 3'----------|AACTGt(-35)|---------|ATaTtA(-10)|----------------------|T|------------5' |転写開始点 |---------------------> mRNA
-
図1.プロモーター配列と転写開始位置の位置関係[2]
-
出現頻度が50%以上の塩基は大文字で、以下のものは小文字にして示す。
- T : ターミネーター
- -10 : -10ボックス
- -35 : -35ボックス
-
出現頻度が50%以上の塩基は大文字で、以下のものは小文字にして示す。
-10ボックス:T(80)A(95)T(45)A(60)A(50)T(96) -45ボックス:T(82)T(84)G(78)A(65)C(54)A(45)
-
図2.-10ボックスと -35ボックスの共通配列および各塩基の存在確率[1]
- 確率の百分率を各構成塩基の右に () 内で記す。
UPエレメント
コアプロモーターエレメントという、-10ボックスも-35ボックスも含む極めて強いプロモーターには、さらに上流にUPエレメントまたは上流要素と呼ばれる特殊な塩基配列を含むものがある。これはαサブユニットに特異的に結合することで、転写開始段階において外れやすいRNAポリメラーゼのDNAとの結合を強化する[5]。たとえば、大腸菌にはrRNAをコードする7個の遺伝子 (rrn gene) があるが、これらにはUPエレメントが存在する。結果、増殖など大量のrRNAが必要なときには7個のrrn遺伝子だけで細胞中の転写の大部分を占める。そのうちの一つrrnB P1遺伝子はUPエレメントにより転写頻度が40倍増強されるという[2]。
-60 -50 -40 -30 -20 -10 +1
| | | | | | |
5’----T[CAGAAAATTATTTTAAATTTC]CTC[TTGTCA]GGCCGGAATAACTCCC[TATAAT]GCGCCACCACT---3'
UPエレメント -45ボックス -10ボックス
図3.rrnB P1 プロモーターの概略 <ref name='weaver140' />
その他の制御装置
rrn geneのプロモーターを制御するのはその塩基配列だけではない。開始NTP (iNTP) とグアノシン5'二リン酸3'二リン酸 (ppGpp) の2つの小さな分子も外部から制御を行う[6]。iNTP存在下は転写産物の材料であるヌクレオチド濃度が高いようだ。INTPは開放型プロモーター複合体を安定化することで転写開始を促す。一方、細胞内のアミノ酸濃度が低く、タンパク質合成がしにくいときはrRNAの合成も必要なくなる。リボソームはアミノ酸を持たないtRNAが結合すると、アミノ酸の不足を感知する。すると、RelAというリボソーム関連タンパク質が警告を受け、ppGppを合成する。警告(alarm)からppGppをアラーモンと呼ぶ。開放型プロモーターをさらに不安定にし、転写を抑制する。
真核生物のプロモーター
真核生物の場合、真正細菌プロモーターの-10領域に相当する、5'-TATAAA-3' の共通配列を持つ領域(TATAボックスあるいはゴールドバーグ・ホグネス・ボックスと呼ばれる)が-25あるいはさらに上流に存在する。転写開始位置はこのTATA ボックスが決定している場合が多い。
この他、-100~-60の範囲に存在する5'-CCAAT-3' の共通配列を持つ領域(CAAT ボックスと呼ばれる)や、-60~-40の範囲に存在する5'-GGCGGG-3' の共通配列を持つ領域(GC ボックスと呼ばれる)がよく知られているが、これらは転写の促進に働いていると考えられている。
真核生物の場合、RNAポリメラーゼには3つの種類があり、Pol I、Pol II、Pol IIIと呼び分けられる。転写開始に必要となる因子、プロモーター領域の配列、転写の様式はそれぞれ異なる。それぞれが担当する遺伝子のプロモーターについて次の各項にて紹介する。
また、同じく転写開始を制御しながら転写開始点からとても離れたエンハンサーもある。プロモーターと同様に機能する配列を持ち、またプロモーターに結合するタンパク質と相互作用する。プロモーターとエンハンサーの違いはむしろ便宜的で、どちらに分類しても差し支えのない配列もある。
クラスIプロモーター
クラスIプロモーターがコードするのはrRNA前駆体だけである。例外として、トリパノソーマという原核生物で発見された2種類の遺伝子はRNAポリメラーゼIによりタンパク質を発現する[7]。そのため、1つのゲノムに何百とあるがその配列は全く同じ。しかし、生物種ごとの多様性は、配列要素をいくつも持つクラスIIプロモーターよりはるかに大きい。保存された配列は転写開始部位を囲むATが豊富なイニシエーター initiator:rINR だけしか確認されていない[注釈 2][7]。
2つの離れた領域からなり、その一つのコアプロモーター core promoter は転写開始点の周辺-45~+20にある[7][8]。rINR近くの配列を除くと、プロモーター中配列としては珍しくGCに富む。もう一つは-180~-107に位置する上流プロモーター配列(upstream promoter element、UPE)である[7][9]。どちらもGCに富む配列により転写効率を左右する[9]。取りうる塩基配列は幅広いものの、2つの配置は多くの真核生物で共通する。これらの領域を発見した錢澤南 Robert Tjian らはリンカースキャン変異導入解析を用いてヒトにおける2つの距離の重要性を証明した[7]。間の塩基配列をわずか16bp取り除いただけで、プロモーター活性は野生型の40%まで低下した。44bpならたったの10%に落ち込んだ。一方、28bp長くしても活性に変化はなかった。49bpまで加えて70%になった。プロモーター効率はDNAの除去に大きな影響を受けるようだ。
クラスIIプロモーター
クラスIIプロモーター class II promoter は2つの配列から構成されている。その一つであるコアプロモーター core promoter はふつう40~60bpほどの大きさである[10]。4つ以下に細分化されており、-33位を中心に存在するTATAボックス TATA box[注釈 4]、そのすぐ上流のTFV IIB認識エレメント TF IIB recognition element:BRE、転写開始部位を中心とするイニシエーター initiator:Inr、さらに下流に下流プロモーターエレメント downstream promoter element:DPE のいくつかを含む[注釈 5]。DPEはさらに細分化され、同名のDPEのほか、下流コア配列 downstream core element:DCE、モチーフ10配列 motif ten element:MTE がある[10]コアプロモーターは基本転写因子と結合し、RNAポリメラーゼに与える[注釈 3]。一方、クラスIIプロモーターのもう一つの要素は上流プロモーターエレメント upstream promoter element:UPE (上流制御配列 upstream control element とも)で、転写開始に関与するほかの転写因子が結合する。次の項で各エレメントを詳しく紹介する。
RNAポリメラーゼIIは伝令RNA(mRNA)前駆体のヘテロ核RNA(hnRNA)と少数の核内低分子RNA(snRNA)を転写する。
BRE TATA Inr DCE I DCE II DPE DCE III
---[(GGG/CCA)CGCCC][TATA(A/T)A(A/T)]---//---[(CC/TT)AN(TCC/ATT)]-[CTTC]----[CTGT]-------[(A/G)G(A/T)CGTG][AGC]---
TF2B TBP TF IID TF IID TF IID TF IID
図4.一般的なクラスIIプロモーター by『ワトソン 遺伝子の分子生物学』p397
上にプロモーターの配列要素の略称を、下に結合する基本転写因子を示す。[]の中は共通配列である。MTEは示さなかったが、
DPEのすぐ上流にある。
TATAボックス
TATAボックスはクラスIIプロモーターの中でもっともよく研究されている。 原核生物の-10ボックスと相同だと考えられているが、10bp上流にある-10ボックスに対して25~30bp上流と、転写開始部位からの距離に大きな違いがある。非鋳型鎖[注釈 6]において共通配列TATAAAを含み、高等真核生物では右端のAが転写開始部位の25~30bp上流にある[11]。通常、この後にさらにAかTが続き、GCに富む配列に囲まれていることが多い[12]。共通配列には多くの例外があり、ウサギのβグロビン遺伝子の場合にはグアニンやシトシンが混ざっている[11]。始まりもTATAではなくCATAだ。また、特化遺伝子[注釈 7]には必ずTATAボックスはあるが、ないプロモーター(TATAレスプロモーター TATA-less promoter )もしばしば存在し、全プロモーターの50%かそれ以上である可能性が示唆されている[12]。これらには次の2種類でよくみられる[11]。
- ハウスキーピング遺伝子
- 細胞の生命維持に必要な生化学的経路を制御するため、事実上すべての細胞で常に活性状態にある。例えば、細胞のヌクレオチド合成に必要なアデニン脱アミノ化酵素やチミジル酸合成酵素、ジヒドロ葉酸還元酵素などおよび、ウイルスの後期タンパク質をコードする。これらの遺伝子はTATAボックスの代わりにGCボックス[11]か、下流プロモーターエレメントがある[12]。実際、ショウジョウバエのクラスIIプロモーターの約70%で下流プロモーターエレメントが務める[11]。ハウスキーピング遺伝子の場合、プロモーターだけでは活性が弱く、さらに上流に転写を促進するアクチベーターが配置されている。
- 発生を制御する遺伝子
- ハエのホメオティック遺伝子や哺乳類の免疫系の発達時に働く遺伝子などである。
TATAボックスの役割は細胞により異なる。 クリストファー・ブノアとピア・シャンボンらの実験では、SV40の初期遺伝子で転写開始部位を正確に決定することを明らかにした[13]。これはTATAボックスからの距離に準拠し、開始部位の配列を本来から変えても転写開始に支障はない。また、この以前の1981年にTATAボックスを欠損させても、起点はバラバラになるが、やはり開始頻度は減少せずにむしろ増えることが確認された[11]。TATAボックスの一部は転写効率を調節しない。
一方で、スティーブン・マックナイトとロバート・キングズバリらはヘルペスウイルスのチミジル酸合成酵素がそのTATAボックスを除去することで大いに転写されにくくなることを発見した[13]。リンカースキャン変異導入解析でプロモーター全体から選んだ10bpを別の配列に置換した結果だ。もっともプロモーター活性が低くなった変異体(LS-29/-18)は、TATAボックス内のGCATATTAがCCGGATCCになったものである[13]。このような遺伝子では発現にTATAボックスは必須である。
イニシエーター
イニシエーターは高効率の転写に必要な、開始部位周辺の-3~+5位に存在する保存配列である[12][注釈 2]。哺乳類の共通配列はPyPyAN(T/A)PyPyで、ショウジョウバエはTCA(G/T)T(T/C)である[14]。ここでPyはピリミジン塩基(CまたはT)、Nは不特定、そして下線付きAが転写開始点を示す。アデノウイルスの主要後期プロモーターに存在するイニシエーターはTATAボックスとともにあり、その下流のあらゆる遺伝子を(非常に弱くだが)活性化させる[14]。また、これは上流プロモーターエレメントやエンハンサーの影響を受けやすい。哺乳類の末端デオキシヌクレオチドトランスフェラーゼ(TdT)遺伝子はTATAボックスも上流プロモーターエレメントもないが、イニシエーターだけでその内部の転写開始部位から基本的な量を発現できる。このことを発表したスティーブン・スメールとデイビッド・ボルチモアはSV40から移植したTATAボックスまたはGCボックスでこの遺伝子の転写を強く増幅できることも発見した[14]。
下流プロモーターエレメント
下流プロモーターエレメント(DPE)はショウジョウバエのゲノムに非常に多い。2,000年にAlan Kutach と James KadonagaはTATAボックスと同頻度に存在することを確認した[14]。これらは転写開始部位より約30 bp下流にあり、共通配列はG(A/T)CGである[15]。
DPEはTATAボックスが欠損したとき、その機能を補う。実際にショウジョウバエでは、DPEがあるTATAボックス欠損プロモーターは数多い。2つは類似点が多く、ともにTF IIDという重要な基本転写因子と結合する。
TF IIB認識エレメント
重要な基本転写因子であるTF IIBが結合するためのプロモーター配列。共通配列は(G/C)(G/C)(G/A)CGCCである[15]。
クラスIIIプロモーター
クラスIIIプロモーター class III promoter は転写開始の過程をその構造により3つに分ける。
- I型。遺伝子内部にあり、Aブロック、中間エレメント、Bブロックを持つもの。TFIIIA、TFIIIB、TFIIIC の三つが必要
- 遺伝子構造の内部にA ブロック(+20 の位置)、B ブロック(+51 から+113 の位置)の二つを持つもの。TFIIIB、TFIIIC の二つが必要。
- TATA ボックス(-25 の位置)、PSE(近位配列要素;-55 の位置)を持つもの。TBP、TFIIIB、PTF の三つの因子が必要
の3つの様式に分かれる。いずれも転写因子の認識配列だけを有するが、3つを次の各項で紹介する。
↓中間エレメント I型 --0--------------========---====----========----------- ボックスA ボックスC II型 --0------------========---------------========--------- ボックスA ボックスB III型 --======----//----========----------========----0------ DSE PSE TATAボックス 図5.3つのクラスIIIプロモーター * 0は転写開始部位
I型
I型 クラスIIIプロモーターとは5S rRNA を発現するプロモーターである。最大の特徴は遺伝子の内部で働くことだろう。ドナルド・ブラウンはケニアツメガエル Xenopus borealis の5S rRNA遺伝子から世界で初めてクラスIIIプロモーターを解析したが、その結果は50~83位上流に存在するという驚くべきものだった。上流から欠失させていくと+55位まで、下流からなら+80位まで転写は影響を受けない[9]。しかし、これを踏み越えると転写は全く起きなくなるのだ。このプロモーターはRNAポリメラーゼIを自身の上流から転写を始めさせる[注釈 8]。その後、ロバート・ローダー Robert Roeder らがボックスA box A、中間エレメント intermediate element、ボックスC box C というI型プロモーターの3領域を発見した[16]。
ボックスAは基本転写因子のTF IIIAと、ボックスCはTF IIICと結合する。これには順番があり、まずTF IIIAから続いてTF IIICが来る。これらは構築因子と言われ、2つがそろうと転写開始点にTF IIIBが結合して転写は始まる。
II型
II型クラスIIIプロモーターは tRNAやVA RNAなどの遺伝子といった、ほとんどのクラスIIIプロモーターに該当する。I型同様遺伝子の内部に存在する。I型の同名領域と酷似するボックスAとボックスBを持つ[17][18]。2つの距離はまちまちだが、普通あまり短すぎると機能を失う[18]。
AボックスにTF IIICが結合することでTF IIIBが転写開始点に結合する。
III型
ほかの2つと異なり、III型は遺伝子上流にある。ヒト7SK RNAやヒトU6 RNAなどの核内低分子RNA (snRNA) を発現させる。1985年にElisabetta Ulluとアラン・ウィーナーが7SL RNA遺伝子から発見したが、そのときの実験では、この領域の除去は転写効率を50~100分の1まで低下させた[17]。ただし、まったく転写されなくなるわけではないので、内部にII型プロモーターも含んでいると考えられる。このことから、ヒトゲノムにおいて何百もある7SL RNA偽遺伝子やそれに関連するAlu配列はあまり転写されない理由が考え出された[17]。これらには高頻度の転写に必要なIII型プロモーターがないのだろう。
III型プロモーターだけを持つ一つの例は7SK RNA遺伝子である。この事実に気づいたマリアルイサ・メッリらは、-37位から下流なら欠損させても転写のレベルを維持できることを証明した[19]。この遺伝子をはじめ、U6 RNA遺伝子やEBER2遺伝子などはII型プロモーター同様にTATAボックスを持つという特徴がある[注釈 9]。実は、TATA結合タンパク質(TBP)はクラスIIプロモーターだけでなく、クラスIとクラスIIIの転写にも関与する。ほかにオクタマー octamer、近位配列因子 proximal sequence element:PSE と呼ばれる配列を含む。転写開始はTATAボックスだけを含む短い領域で行われるが、2つの配列が加わると転写効率は劇的に増す[20]。
注釈
- ^ a b 転写並びDNA複製は鋳型鎖で3’から5’末端への方向に進む。よって、生物学者は鋳型鎖の5’末端側を上流 upstream、3’末端側を下流 downstream と呼ぶのが通例だ。1個のデオキシヌクレオチド(DNAの構成単位)の相対的な位置を表す際、間のデオキシヌクレオチドの数に、そこから上流なら-、下流なら+を前に付けて表記する。プロモーターの中には最初のリボヌクレオチドが塩基対形成をする転写開始点 startpoint があり、転写に関わるほかの遺伝子の位置を示す場合に『+1』とする。0と番号付けるデオキシヌクレオチドはなく、下流の1個前は-1だ。
- ^ a b c DNAの塩基配列の中には、重要な意味を持つために同種間または異種間にわたって存在するものがある。これを保存された conserved と形容する。
- ^ a b プロモーターに結合したRNAポリメラーゼはすぐに転写を実行するわけではなく、その前準備となる開始段階を必要とする。その一環は基本転写因子と呼ばれる多数のタンパク質とともに転写開始前複合体を成すことだ。転写開始段階はさらにいくつかの手順を追うが、その各段階で複合体は変化する。
- ^ TATAボックスの位置は出芽酵母ではまちまちで、中には300bp以上も上流に離れていることもある。
- ^ クラスIIプロモーターはコアプロモーターと上流プロモーターエレメントを合わせた5つから構成されるが、天然で全て揃っていることは稀である。プロモーターに幅広い特性を与えるためか、一つ以上が欠損している場合が多い。例えば酵母に関する最近の研究においては、TATA-containing core promoter(TATA ボックスを含むコアプロモーター)は、わずか約19% であったと報告されている。
- ^ 核内の通常のDNAは二本で一本の二重らせんを形成しており、転写されるのはこのうち一本だけだ。よって、されるほうを鋳型鎖、されないほうを非鋳型鎖と呼び分ける。
- ^ 特化遺伝子とは、特定の細胞でしか発現しない遺伝子だ。皮膚細胞のケラチンや赤血球のヘモグロビンなどをコードする。
- ^ ケニアツメガエルで発見された5S RNA遺伝子のクラスIプロモーターは、組み込めばどんなDNAでもその約55bp上流で転写を開始させる。本来の遺伝子にも当然この機能は働くが、本来の転写開始点を欠損させて試すとやはり約55bp上流に一番近いプリン塩基から始まる。
- ^ これらの遺伝子がポリメラーゼIIではなくポリメラーゼIIIによって転写されることを証明する方法は、ポリメラーゼIIを阻害するα-アマニチンを用いることだ。ポリメラーゼIIで転写されるならαアマニチンは低濃度で十分だが、高濃度が必要となるならばそうではないに違いない。阻害様式から関与するのがポリメラーゼIIIであることを証明できる。
出典
- ^ a b c d e Lewin 2006, p. 233
- ^ a b c d e Weaver 2008, p. 140
- ^ Watson 2010, p. 382
- ^ a b c Lewin 2006, p. 234
- ^ a b Watson 2010, p. 384
- ^ Weaver 2008, p. 141
- ^ a b c d e Weaver 2008, p. 293
- ^ Lewin 2006, p. 539
- ^ a b c Lewin 2006, p. 540
- ^ a b Watson 2010, p. 397
- ^ a b c d e f Weaver 2008, p. 289
- ^ a b c d Lewin 2006, p. 543
- ^ a b c Weaver 2008, p. 290
- ^ a b c d Weaver 2008, p. 291
- ^ a b Weaver 2008, p. 292
- ^ Weaver 2008, p. 294
- ^ a b c Weaver 2008, p. 295
- ^ a b Lewin 2006, p. 541
- ^ Weaver 2008, p. 296
- ^ Lewin 2006, p. 542
参考文献
- Benjamin Lewin著、菊池韶彦訳 『遺伝子』(第8版)東京化学同人、2006年。ISBN 978-4807906307。
- Robert F. Weaver (著), 杉山弘 (監訳), 井上丹 (監訳), 森井孝 (監訳) 『ウィーバー 分子生物学』(第4版)化学同人、2008年。 ISBN 978-4759811568。
- James D. Watsonほか、監訳者:中村桂子 『ワトソン 遺伝子の分子生物学』(第6版)東京電機大学出版局、2010年。 ISBN 978-4501625702。
関連項目
外部リンク


| 国立図書館 | |
|---|---|
| その他 | |
プロモーター
出典: フリー百科事典『ウィキペディア(Wikipedia)』 (2022/05/08 07:18 UTC 版)
イニシエーターとペアを組んで戦う民警社員。座学と実技の試験を受け民警許可証(ライセンス)を取得することで就くことができる。イニシエーターの指導監督、および戦闘時における指揮が主な役割である。プロモーターはイニシエーターと違い一般の人間であるもの、バラニウム製の重銃器や近接武器を持ち、戦闘中において主に後衛を務める。その理由はイニシエーターは強い治癒再生力を持つので急所以外の誤射は致命傷にならないこと、比較的安全な場所から指示が出せること、万一イニシエーターが不審な行動をとった場合即刻殺すことができる位置取りをする必要があること、である(3つ目はプロモーターの精神的安定を保つ意味合いが強い)。このことから基本的に戦闘はイニシエーターが担当といえるが、実際のところ民警となる人間はチンピラ上がりなど好戦的な人物が多く、指示を出すどころか自ら突っ込んでイニシエーターがその補助をするというパターンも見受けられる。イニシエーターを道具扱いする者、家族のように接する者、関係は十人十色である。
※この「プロモーター」の解説は、「ブラック・ブレット」の解説の一部です。
「プロモーター」を含む「ブラック・ブレット」の記事については、「ブラック・ブレット」の概要を参照ください。
「プロモーター」の例文・使い方・用例・文例
- プロモーターのページへのリンク




