よう‐みゃく〔エフ‐〕【葉脈】
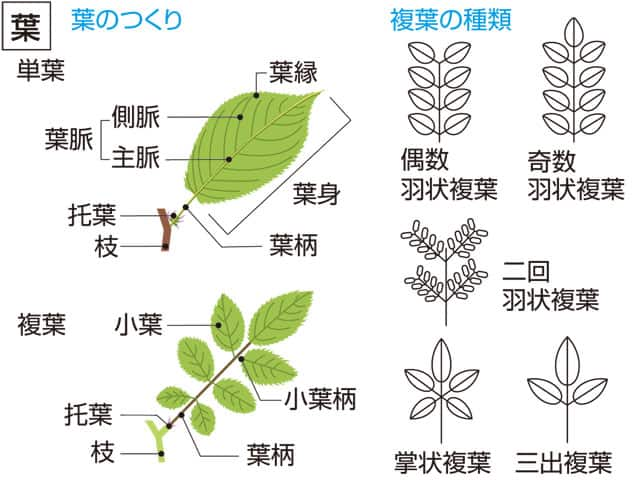

葉脈
出典: フリー百科事典『ウィキペディア(Wikipedia)』 (2023/12/01 17:48 UTC 版)

葉脈(ようみゃく、英: vein, nerve)とは、葉における維管束であり、葉の表面に筋として見える構造である(図1)。葉脈は茎の維管束につながり、水や栄養分を葉に行き渡らせ、葉での光合成によってつくられた有機物を葉から運ぶ通路となる。また葉脈は細胞壁が硬化した細胞を含み、葉の機械的支持にも機能している。
1枚の葉に複数の葉脈がある場合、最も太い葉脈は主脈(または一次脈)、そこから分岐した太い葉脈は側脈(または二次脈)とよばれる。主脈が中央脈となっていることも多い。また被子植物では、細脈によって葉身が細かく区画化されている(図1)。葉脈の配列様式には多様性があり、そのパターンは脈系(venation)とよばれる。脈系は、一般的に網状脈、平行脈、二又脈、単一脈に分けられる。
構造
葉脈は、葉の葉身において維管束とそれを取り囲む組織からなる構造であり、外形としては葉の表面の筋として見える[1][2][3][4][5]。葉脈の維管束は木部と師部からなり、木部を通じて根から輸送されてきた水や無機栄養分を葉に行き渡らせ、また葉の光合成産物を師部に積み込んで(ローディング)他の器官へ輸送する[5][4]。さらに葉脈を構成する維管束にはリグニン化した細胞が含まれ、加えて厚壁組織や厚角組織を伴うことがあり、葉身の機械的支持の役割も担う[5]。太い葉脈では、維管束が発達した組織に囲まれて葉の裏側(背軸側)に突出した肋になっていることが多い[1]。
外部形態
1枚の葉の中に太さが異なる複数の葉脈がある場合、明らかに最も太い葉脈(複数の場合もある)を主脈(main vein)という[1][2]。多くの場合、主脈は葉の中軸にある中央脈(中脈、central vein)でもある[1][2](下図2a)。ただしヤツデのように掌状に分裂した葉では、複数の主脈が掌状に伸びているが、明瞭な中央脈はない[2]。またイチョウの葉のように、主脈も中央脈も明瞭ではないこともある。中央脈を含む部分が線状に隆起しているものは、中肋(midrib, costa[注 1])ともよばれる[1][2]。コケ植物の蘚類の葉も中肋をもつことがあるが、コケ植物は維管束をもたず、葉脈はない。
主脈から分岐した太い葉脈は側脈(lateral vein)とよばれ、分岐順によって一次側脈(primary lateral vein)、二次側脈(secondry lateral vein)、のようによばれる[1][2](下図2a)。主脈を一次脈(primary vein)、一次側脈を二次脈(secondary vein)、二次側脈を三次脈(tertiary vein)、のようによぶこともある[1][2][3]。また、三次脈以降を一括して二次間脈(intersecondary vein)とよぶことがあるが、細い側脈を二次間脈とよぶこともある[1][3]。比較的大きな葉脈を大脈(major vein)、小さな葉脈を小脈(minor vein)とよんでいることもある[1]。


主脈や側脈から生じてこれらの間をつないだり、網目をつくったり、末端が遊離している細い葉脈は、細脈(veinlet)とよばれる[1][2][3](上図2a, b)。細脈によって形成される末端の網目は最終区画(ultimate areole)、最終区画の内外に遊離する葉脈は脈端(遊離端、盲管、vein ending)とよばれる[1][2][6][3](上図2b)。
内部構造
茎の維管束から分岐して葉に入る維管束は、葉跡(ようせき; leaf trace)とよばれる[7][8][9]。小葉植物以外では、茎の維管束において、葉跡が別れた際にその上側に維管束がない隙間が生じ、この隙間は葉隙(ようげき; leaf gap)とよばれる[7][8]。1枚の葉に入る葉跡の数、およびそれに対応する葉隙の数には、多様性がある[7][10]。
種子植物の茎の維管束は、基本的に内側が木部、外側が師部の並立維管束であるため、これが葉に入った葉脈では、上側(向軸側)に木部、下側(背軸側)に師部が配置している[1][8][11](下図3a, b)。また原生木部(最初に分化する木部)は上側、原生師部(最初に分化する師部)は下側に位置する[8]。ただし、木部の上下に師部が位置している複並立維管束をもつものもある[1][12]。また大葉シダ植物では、木部が師部で囲まれている包囲維管束をしばしばもつ[13]。
太い葉脈では上記のように木部と師部がセットになっているが、細い葉脈や脈端ではしばしば木部の道管がなく1–2本の仮道管のみになっており、また明瞭な師部を欠いて師管の伴細胞にあたる細胞が転送細胞(transfer cell)となっていることがある[1][11][8]。転送細胞では細胞壁が入り組んで表面積が広くなっており、効率的な物質転送に機能すると考えられている[8]。


葉脈を構成する維管束は、しばしば柔細胞からなる細胞層である維管束鞘(bundle sheath)で包まれている[7][11][8][12][14](上図3a)。維管束鞘は、通道や一時的な貯蔵に機能していると考えられている[14]。維管束鞘の位置や発生は、内皮に似ている[8]。
また、維管束鞘から上側(向軸側)、下側(背軸側)、または上下両側の表皮方向に柔組織が発達していることがあり、維管束鞘延長部(bundle sheath extension)とよばれる[7][11][8][12][14]。維管束鞘延長部は、維管束と表皮の間の物質移動に関わっていると考えられている[11][12]。またこのような維管束鞘延長部の存在によって葉肉が区画化されることになり、葉身の一部に起こった傷害が、他の部分に波及することを防止するとも考えられている[8]。また一次脈や二次脈など太い葉脈では、柔組織や厚壁組織からなる維管束鞘延長部が発達して葉脈部が突出して肋となることがある[8](上図3a, b)。ただしこのような発達した構造は、狭義の維管束鞘延長部には含めないこともある[8]。
トウモロコシなどC4型光合成を行う植物では、維管束鞘(またはその一部)は葉緑体を多く含む細胞から構成され、さらにこれが葉肉細胞で取り囲まれている[7][11][15][16](上図3b)。このような維管束を囲む構造はクランツ構造(Kranz anatomy)[注 2]とよばれ、リンゴ酸などに固定された二酸化炭素が放出、再固定される場となる[15]。このような維管束鞘細胞の細胞壁にはふつうスベリンなどが沈着して二酸化炭素の拡散を防止し、そのため維管束鞘細胞の二酸化炭素濃度は大気の4.5–18倍に達することが報告されている[17]。
葉脈は、シュート頂分裂組織に由来する前形成層から分化する[1][9]。種子植物では、葉原基の前形成層から形成された維管束が基部側へ(求基的に)成長し、茎にあるより古い維管束系と接続する[9][18]。シロイヌナズナでは、葉縁部にある特定の箇所でオーキシン合成が起こり、PINオーキシン排出キャリアタンパク質などによってオーキシンの流路が形成され(カナリゼーション canalization)、これに沿って維管束(葉脈)が形成されることが報告されている[9]。
脈系
葉脈の配列様式のことは、脈系(脈理)とよばれる[2][4][3][6][19][20]。脈系は以下のように幾つかの型に類別される。ただしそれぞれの区分ははっきりしているわけではなく、中間的なものもある。
網状脈
主脈、側脈、細脈が結合して網目を形成する脈系は、網状脈系(網状脈、reticulate venation, netted venation)とよばれる[2][6][19][3][20]。主脈や側脈の配列によって、以下のような型に分けられる。ただし中間的なものもある。
-
- 羽状脈系(羽状脈、pinnate venation)[2][4][19][3][20]
- 一次側脈が羽根状に配列したもの(下図4a, b)。被子植物で最も一般的な脈系であり、ケヤキ(ニレ科)、ハンノキ(カバノキ科)、イチイガシ(ブナ科)、チドリノキ(ムクロジ科)、ヤマボウシ(ミズキ科)などに見られる[2]。一次側脈(二次脈)が葉縁に達するものは縁終脈型(craspedodromous; 下図4a)、一次側脈が葉縁に達せず弧となるものは輪弧型(camptodromous; 下図4b)とよばれる[12]。
- 掌状脈系(掌状脈、palmate venation)[2][4][19][3][20]
- 複数の主脈が1点から広がる掌状に配列したもの。主脈が3本あるものは三行脈とよばれ(下図4c)、クスノキ(クスノキ科)、サルトリイバラ(サルトリイバラ科)、カラスウリ(ウリ科)、カクレミノ(ウコギ科)、カンボク(ガマズミ科)などに見られる[2]。主脈数がそれより多いものは多行脈(下図4d)とよばれ、カツラ(カツラ科)、カナムグラ(アサ科)、イロハモミジ(ムクロジ科)などに見られる[2]。また、最も外側の主脈の基部付近から太く長い一次側脈が伸びているものは、鳥足状脈系(鳥足状脈、pedate venation)ともよばれ、ウマノアシガタ(キンポウゲ科)やスズカケノキ(スズカケノキ科)などに見られる[2][3][20](下図4e)。





平行脈
多数の葉脈(一次脈、主脈)が分岐することなく平行にならんでいる脈系は、平行脈系(平行脈、parallel venation)とよばれる[2][6][19][3][20][21](下図5a–d)。ふつう葉の先端ではすべての一次脈が収束して厳密には平行ではないため、条線脈(striate venation)とよばれることもある[2][12]。平行にならんだ一次脈の間はふつう細い葉脈(結合脈)でつながっており、網目を形成する点では網状脈と変わらない(ただし網目はふつう方形)[6][19]。イネ科やアヤメ科など単子葉類に一般的に見られる。オオバコ(オオバコ科)などに見られる脈系は、網状脈とされることも[2]、平行脈とされることもある[21][20](下図5e)。





二又脈
葉脈が二又に分岐して網目をつくらない脈系は、二又脈系(二又脈、二叉脈、叉状脈、dichotomous venation)とよばれる[2][19][3][20][22]。シダ類の多くは二又脈をもち(下図6a)、ソテツ類(一部を除く[23])、イチョウ、ナギ、ナンヨウスギ属などの裸子植物にも見られる[2][6](下図6b, c)。被子植物では、キルカエアステル科で報告されている[2][6][12](下図6d)。




シダ類の中でも、コウヤワラビなどは網状脈をもつ[6]。また裸子植物の中で、典型的な網状脈はグネツム属のみに見られる[6]。しかしその発生過程を調査した研究からは、これらの網状脈は二又脈の連結によって形成されることが示されている[6]。
単一脈
中央脈1本のみがあって分岐しない脈系は、単一脈系(simple venation)とよばれる[2]。小葉植物(ヒカゲノカズラ植物; 下図7a)、トクサ類、トウヒやイチイなど多くの球果類(針葉樹; 下図7b, c)、ガンコウランやツガザクラ(ツツジ科; 下図7d)などに見られる[2][20]。




進化
維管束植物の中で、小葉植物(ヒカゲノカズラ類)、大葉シダ植物、種子植物は独立に葉を獲得したと考えられており、茎から葉に伸びる維管束である葉脈も独立に生じた[24][25][26]。葉の維管束は、小葉植物や大葉シダ植物では基部側から先端側へ(求頂的)分化するのに対して、種子植物では先端側から基部側へ(求基的)分化することが知られている[9][25]。
シダ植物や裸子植物では、葉脈はふつう単一脈や二又脈であり、脈系は単純である[5][18]。一方、被子植物は、網状脈や平行脈などの階層性を示す複雑な葉脈をもつことが多い[5][18]。特に細脈が発達して葉脈の総延長の約90%以上を占めることもある[5]。葉脈密度は、葉の通水コンダクタンスや光合成速度と強い正の相関があることが示されており、細脈が高密度に配置されていることで、効率的な光合成が可能になっていると考えられている[5]。白亜紀に被子植物が急速に多様化・優占した理由の一つとして、この時期に低下した二酸化炭素濃度に対して被子植物の高い葉脈密度が有利に働いたことが示唆されている[27]。
葉脈標本

葉脈にはリグニンなどが沈着した固い細胞壁をもつ細胞が含まれるため、葉の他の部分より残りやすい。葉脈だけが残った葉は葉脈標本(スケルトンリーフ、透かし葉)ともよばれ、しおりなどに利用されることがある[28][29][30][31][32]。人工的には、アルカリ水溶液によって葉肉部を溶解させて除去することで作成され、漂白して着色することもある[28][29][30][31][32](図8)。
ギャラリー
-

-
 キササゲ属(ノウゼンカズラ科)の葉脈
キササゲ属(ノウゼンカズラ科)の葉脈 -
 カロデンドラム・カペンセ(ミカン科)の葉脈
カロデンドラム・カペンセ(ミカン科)の葉脈 -

-

-
 イチジク(クワ科)の葉脈
イチジク(クワ科)の葉脈 -

-

-

-

-
 薄嚢シダ類の葉(二又脈)
薄嚢シダ類の葉(二又脈) -













脚注
注釈
出典
- ^ a b c d e f g h i j k l m n o 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “葉脈”. 岩波 生物学辞典 第5版. 岩波書店. p. 1427. ISBN 978-4000803144
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z 清水建美 (2001). “葉脈と脈系”. 図説 植物用語事典. 八坂書房. pp. 134–140. ISBN 978-4896944792
- ^ a b c d e f g h i j k l 原襄 (1994). “葉”. 植物形態学. 朝倉書店. pp. 36–46. ISBN 978-4254170863
- ^ a b c d e 「葉脈」『日本大百科全書(ニッポニカ)』。コトバンクより2023年6月7日閲覧。
- ^ a b c d e f g “葉脈”. 光合成事典(Web版). 日本光合成学会 (2020年5月12日). 2023年6月7日閲覧。
- ^ a b c d e f g h i j 熊沢正夫 (1979). “18.7 脈理”. 植物器官学. 裳華房. pp. 215−219. ISBN 978-4785358068
- ^ a b c d e f 清水建美 (2001). “内部形態”. 図説 植物用語事典. 八坂書房. pp. 157–164. ISBN 978-4896944792
- ^ a b c d e f g h i j k l 原襄 (1994). “葉の構造”. 植物形態学. 朝倉書店. pp. 59–68. ISBN 978-4254170863
- ^ a b c d e L. テイツ, E. ザイガー, I.M. モーラー & A. マーフィー (編) (2017). “葉の脈理パターン”. 植物生理学・発生学 原著第6版. 講談社. pp. 565–572. ISBN 978-4061538962
- ^ 田村道夫 (1999). “節の構造”. 植物の系統. 文一総合出版. pp. 144–145. ISBN 978-4829921265
- ^ a b c d e f 原襄 (1972). “8.5 葉の構造”. 基礎生物学選書 3. 植物の形態. 裳華房. pp. 178–190. ISBN 978-4-7853-5103-8
- ^ a b c d e f g アーネスト・ギフォード & エイドリアンス・フォスター (著) 長谷部光泰, 鈴木武 & 植田邦彦 (監訳) (2002). “脈系、単子葉植物と双子葉植物の脈系、被子植物における脈系の系統発生、葉の組織”. 維管束植物の形態と進化. 文一総合出版. pp. 495–503. ISBN 978-4829921609
- ^ アーネスト・ギフォード & エイドリアンス・フォスター (著) 長谷部光泰, 鈴木武 & 植田邦彦 (監訳) (2002). “葉の内部構造”. 維管束植物の形態と進化. 文一総合出版. pp. 257, 260–265. ISBN 978-4829921609
- ^ a b c 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “維管束鞘”. 岩波 生物学辞典 第5版. 岩波書店. p. 59. ISBN 978-4000803144
- ^ a b c L. テイツ, E. ザイガー, I.M. モーラー & A. マーフィー (編) (2017). “無機炭素濃縮機構:C4炭素回路”. 植物生理学・発生学 原著第6版. 講談社. pp. 220–228. ISBN 978-4061538962
- ^ 吉村泰幸. “C4植物の葉の構造いろいろ”. C4 plants Party. 農研機構 農業環境変動研究センター. 2023年6月16日閲覧。
- ^ 吉村泰幸. “C3植物との違い”. C4 plants Party. 農研機構 農業環境変動研究センター. 2023年6月16日閲覧。
- ^ a b c 長谷部光泰 (2020). “16.2 高次網状脈系の進化”. 陸上植物の形態と進化. 裳華房. pp. 216–217. ISBN 978-4785358716
- ^ a b c d e f g 原襄 (1972). “8.3 葉脈とその分化”. 基礎生物学選書 3. 植物の形態. 裳華房. pp. 169–176. ISBN 978-4-7853-5103-8
- ^ a b c d e f g h i 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “脈系”. 岩波 生物学辞典 第5版. 岩波書店. p. 1363. ISBN 978-4000803144
- ^ a b 「平行脈」『日本大百科全書(ニッポニカ)』。コトバンクより2023年6月10日閲覧。
- ^ 「二又脈」『日本大百科全書(ニッポニカ)』。コトバンクより2023年6月9日閲覧。
- ^ アーネスト・ギフォード & エイドリアンス・フォスター (著) 長谷部光泰, 鈴木武 & 植田邦彦 (監訳) (2002). “葉”. 維管束植物の形態と進化. 文一総合出版. pp. 366–370. ISBN 978-4829921609
- ^ 長谷部光泰 (2020). “8.3 小葉”. 陸上植物の形態と進化. 裳華房. pp. 128–129. ISBN 978-4785358716
- ^ a b 長谷部光泰 (2020). “9.2 枝から葉への進化:現代的テローム説”. 陸上植物の形態と進化. 裳華房. pp. 144–148. ISBN 978-4785358716
- ^ 長谷部光泰 (2020). “11.3 葉の進化”. 陸上植物の形態と進化. 裳華房. p. 176. ISBN 978-4785358716
- ^ De Boer, H. J., Eppinga, M. B., Wassen, M. J. & Dekker, S. C. (2012). “A critical transition in leaf evolution facilitated the Cretaceous angiosperm revolution”. Nature Communications 3 (1): 1221. doi:10.1038/ncomms2217.
- ^ a b “作ってみよう!葉脈標本~スケルトンリーフ~”. Youtube. 環境情報チャンネル【千葉県環境研究センター】. 2023年6月7日閲覧。
- ^ a b “葉脈標本をつくろう”. さいたま市 (2016年6月16日). 2023年6月7日閲覧。
- ^ a b 小川誠. “葉脈標本できれいなしおりづくり”. 徳島県立博物館. 2023年6月8日閲覧。
- ^ a b マリ (2021年8月4日). “キャンプの思い出や夏休みの工作にも!きれいな葉っぱで葉脈標本を作ってみよう”. BE-PAL. 小学館. 2023年6月7日閲覧。
- ^ a b 神山真由美. “葉の中に隠れた繊細なアート。子どもと楽しむスケルトンリーフ(葉脈標本)づくり”. GardenStory. 2023年6月9日閲覧。
関連項目
外部リンク
- 「葉脈」。コトバンクより2023年6月7日閲覧。
- “葉脈”. 光合成事典(Web版). 日本光合成学会 (2020年5月12日). 2023年6月7日閲覧。
- “葉のはたらき(気孔と葉脈)”. NHK for School. 2023年6月7日閲覧。
- “葉脈標本データベース”. 国立科学博物館. 2023年6月10日閲覧。
- Ellis, B., Daly, D. C., Hickey, L. J., Johnson, K. R., Mitchell, J. D., Wilf, P. & Wing, S. L. (2009). Manual of leaf architecture. Cornell University. ISBN 080147518X (英語)
- “フラボンの山野草と高山植物の世界”. 2023年6月10日閲覧。
- “作ってみよう!葉脈標本~スケルトンリーフ~”. YouTube. 環境情報チャンネル【千葉県環境研究センター】. 2023年6月7日閲覧。

葉脈
出典: フリー百科事典『ウィキペディア(Wikipedia)』 (2022/06/09 14:41 UTC 版)
詳細は「葉脈」を参照 葉にはたくさんの筋が入っているのが普通である。これは、茎から葉に入った維管束であり、葉脈と呼ばれる。普通は葉柄から葉の先端へと中心になる脈(主脈、中肋とも)が走り、その途中で側方へと枝の脈(側脈)が伸びる。側脈からはまた枝が出て、それぞれの先でぶつかり、全体として鳥の羽の筋のような(羽状)、あちこちで網目を作る(網状脈)形になることが多い。 単子葉植物では、主脈と側脈が共に葉の根元から先端へと向かい(平行脈)、それらをつなぐ細い脈が横に走る形になるものが多い。 シダ植物門やイチョウでは、葉脈は二又分枝的なものが多い。特にイチョウでは、葉脈はほぼ完全に二又分枝の繰り返しからなる。これは原始的特徴と考えられている。 葉脈の断面を見ると、葉の表側には道管を中心とする木部が、裏面側に師管を中心とする師部がある。これは、葉の表側が茎の中心を向いていること、茎では木分が中心側にあることを考えれば、当然の配置である。 茎の維管束が葉へと入るため、葉の上側では茎の維管束に隙間が生じる。これを葉隙という。
※この「葉脈」の解説は、「葉」の解説の一部です。
「葉脈」を含む「葉」の記事については、「葉」の概要を参照ください。
「葉脈」の例文・使い方・用例・文例
- (花について)葉脈に似た紫を見せる
- 植物の葉脈に関連するさま
- 葉の葉脈の間または昆虫の翅脈の間の部分のような組織またはある部分の小さな空間
- 植物か動物の肋骨状の部分(葉の真ん中の葉脈や、昆虫の翅の肥厚した翅脈のような)
- 1枚の子葉と葉脈が平行に走る葉を持つ胚をつくる種子植物からなる:イネ、ユリ、ヤシ、ランを含む
- 多く枝分かれした高木状のあるいは低木状のサボテンで、明らかな葉脈と円くなった麺状のとげを持ち、通常白の夜咲きの花をつける
- 紫の葉脈がある葉が根元から出てロゼットをつくるミヤマコウゾリナ属の多年草
- 特に金属系の白または金色の葉脈をもつビロードのような葉のために育てられるいくつかの繊細なアジアランのどれか
- 暗色の葉脈のある黄色がかった褐色の花を持つラン
- 各小花に6か8の雄ずい、1年で成熟するどんぐりと葉の端を超えて決して伸びない葉脈を持つ多くの旧世界とアメリカのオークのどれか
- 各小房に4つの雄しべを持つ多くアメリカオークの総称で、ドングリは成熟までに2年を要し、葉脈は通常葉の縁までのびて先端部を形成しているか生い茂っている
- 熱帯常緑の高木と低木の科で、厚みと輝きのある、葉脈の平行した葉を持つ
- 柔らかく軟毛で覆われた葉と茎と、茶色から紫色の葉脈のあるはっきりした黄色い花を有する北アメリカ東部のスミレ
- 葉脈のある長細い葉を持つ、ヨーロッパ原産のオオバコで、広く温帯地域に定着している
- 白がかった紫色の葉脈がある花をつけるアカンサスでヨーロッパ南部で広く栽培されている
- 葉の中央にある葉脈
- 胞子嚢を持つ遊離性葉脈のある熱帯着生シダ
- 叉状に分岐した葉脈
- 側脈という,葉脈の主脈から側方へ分岐した脈
- 植物の葉脈としての平行脈
- >> 「葉脈」を含む用語の索引
- 葉脈のページへのリンク


