節足動物
出典: フリー百科事典『ウィキペディア(Wikipedia)』 (2024/02/12 08:12 UTC 版)
分類
系統関係と体節の相同性
現生の節足動物は、鋏角類(Chelicerata, クモ、サソリ、カブトガニなど)・多足類(Myriapoda, ムカデ、ヤスデなど)・甲殻類(Crustacea, カニとエビ、フジツボ、ミジンコなど)・六脚類(Hexapoda, 昆虫、トビムシなど)という4つの亜門に分類されている。化石種まで範囲を広げれば、三葉虫などを含んだ Artiopoda という過去の亜門や、前述の亜門には明確に収まらない分類群も数多く知られている(後述参照)[4][115]。
それそれの分類群で特に注目される特徴は、先頭複数体節の融合でできた合体節(頭部および前体)の付属肢である。これは前述のような高次分類群によって異なり、例えば各亜門では次のような既形をもつ[6][14][57]:
- 六脚類:触角1対・大顎1対・小顎1対・下唇1枚(1対の小顎から癒合した部分)
- 多足類:触角1対・大顎1対・小顎2対
- 甲殻類:触角2対・大顎1対・小顎2対
- 鋏角類(真鋏角類):鋏角1対・触肢1対・脚4対
- 鋏角類(ウミグモ類):鋏肢1対・触肢1対・担卵肢1対・脚1対
- Artiopoda類:触角1対・脚3対以上
これらの付属肢の由来と対応関係(相同性)は、節足動物の各分類群の系統関係を示唆する重要な指標の1つであるが、分類群ごとの形態の違いや化石群の証拠の欠如により難解な場合が多く、この問題は「Arthropod head problem」として知られている[14][4]。それに対しては「終わりのない論争」(endless dispute)とも呼ばれるほど、昔今を通じて文献記載により様々な解釈を与えられた[119][14]。




21世紀以前の古典的な見解では、鋏角類は中大脳性な触角を退化して鋏角は後大脳性と考えられ[120]、頭部付属肢や呼吸器の類似を基に多足類と六脚類は近縁という説が主流であり[16][121]、付属肢の単枝型と二叉型の基本形態を基に、節足動物を多系統群として2つに分けるという異説すらあった。しかしこれらの知見は、後に発生学・神経解剖学・遺伝子発現・分子系統学など多方面の情報により根本から否定され、節足動物は疑いなく単系統群・鋏角類の鋏角は他の節足動物の第1触角と同様に中大脳性[122][123][124][125]・六脚類は甲殻類に近縁で側系統群の甲殻類から分岐したことが強く示唆される[126][127][128][129][130][131][132]。また、通常では鋏角類とされるウミグモ類の系統位置がしばしば疑問視されるようになり、分子系統解析では鋏角類であることを支持するものが多い[133][134][115]が、2000年代ではそれ以外の節足動物と対置すべき説もあった[135][129][136]。
三葉虫やメガケイラ類をはじめとして、幾つかの化石節足動物の分類群と現生分類群の類縁関係については、未だに定説がない[4]。例えば三葉虫などを含む Artiopoda類は、鋏角類に類縁という伝統的な系統仮説はあった[120]が、頭部構造の類似に基づいて触角をもつ現生群(多足類・甲殻類・六脚類)に近い[137][138][139]、またはどの現生群よりも基盤的[140]とも考えられる[4]。メガケイラ類は先頭の付属肢と脳の類似に基づいて鋏角類に近いとされる[141]が、どの現生群よりも基盤的ともされる[20][14][4]。Hymenocarina類は一時では単調な頭部をもつと解釈され、それを基にどの現生群よりも基盤的と考えられていた[20][14]が、大顎をもつ口器の発見により、多足類・甲殻類・六脚類と同じ系統群に含める説が主流になりつつある[140][142][4][115]。
こうした研究の発展に伴い、節足動物の高次系統に対して様々な系統仮説が提唱されており、以下の例が挙げられる[143][144]。
- 大顎類 Mandibulata
- 現生群では多足類・甲殻類・六脚類からなる。大顎が共有形質とされる。
- 汎甲殻類 Pancrustacea(=八分錘類 Tetraconata)
- 現生群では甲殻類と六脚類からなる。複眼の八分割される硝子体などが共有形質とされる。
- 多足鋏角類 Myriochelata(=矛盾足類 Paradoxopoda)
- 多足類と鋏角類からなる。
- 裂肢類 Schizoramia(='CCT' clade)
- 甲殻類・鋏角類・Artiopoda類などからなる。ニ叉型付属肢が共有形質とされる[145]。
- 無角類 Atelocerata(=気門類 Tracheata、狭義の単肢類 Uniramia sensu stricto)
- 現生群では多足類と六脚類からなる。後大脳性/第2体節付属肢の欠如・気門などが共有形質とされる。
- 広義の単肢類 Uniramia sensu lato
- 有爪動物・多足類・六脚類からなる。単枝型付属肢が共有形質とされる。またこの系統仮説に従えば、有爪動物は節足動物に含まれ、もしくは節足動物が有爪動物に対して多系統群になる[16]。
- Arachnomorpha(=蛛形様類 Arachnata[146])
- Artiopoda類と鋏角類からなる。
- Antennulata
- Artiopoda類・多足類・甲殻類・六脚類などからなる。中大脳性/第1体節由来の第1触角が共有形質とされる。
- 板肢類 Lamellipedia[145][146]
- Artiopoda類と Marrellomorpha類からなる。
2010年代以降では、少なくとも大顎類説、汎甲殻類説、およびウミグモ類が鋏角類に含める説が広く認められる[4][115]。
| 節足動物の内部系統関係 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 節足動物の各亜門(太字)の類縁関係とそれぞれの主要な内部高次系統までの分岐図[4][115]。青枠以内の分類群、すなわち六脚類以外の汎甲殻類は側系統の甲殻類に属する。絶滅群は「†」、類縁関係が議論的なもの(鋏角類とArtiopoda類と大顎類、貝形虫類とヒゲエビ類とウオヤドリエビ類、軟甲類と鞘甲類とカイアシ類)は3本以上の多分岐、単系統性が議論的なもの(節口類、クモガタ類、内顎類)は二重線で示される。亜門が未定・不確実、もしくはそのクラウングループに含まれない絶滅群、およびほとんどの系統解析に含まれないヒメヤドリエビ類はここに示さない。 |
また、現生節足動物の胚発生において、眼と前大脳が由来する先頭の体節、いわゆる先節には付属肢を持たないのが従来の解釈であった。しかし、多くの節足動物の口の前にある蓋状の構造体、いわゆる上唇やハイポストーマは、著しく融合・退化した先節由来の1対の付属肢であることが2000年代以降から有力視されつつある[18][19]。この構造体は、有爪動物の触角や基盤的な節足動物(ラディオドンタ類など)の前部付属肢との相同性まで議論をなされ、初期の節足動物、果ては汎節足動物全般における頭部の起源と進化を示唆する手掛かりの一つとして注目される[113][20][14]。

下位分類

節足動物は記載された種数の最も多い動物門である[1][5][6]。その数は動物に限らず、真核生物の種の大部分をも占める[5][147]。2011年まででは、100万種以上の六脚類・11万種以上の鋏角類・6万種以上の甲殻類・1万種以上の多足類という計120万種以上の現生節足動物が記載されている[148][6]。また、絶滅した三葉虫も大きなグループであり、1万種以上が記載される[149]。
節足動物の主要な高次分類群(綱/亜綱/目)までの下位分類は次の通り。なお、これらは必ずしも2010年代後期以降の主流な系統関係を反映しているとは限らず(例えば甲殻類は六脚類に対して側系統)、分類階級が文献により異なる(鞘甲類・カイアシ類など)・単系統性に議論が続いているものもある(節口類・クモガタ類・六幼生類・内顎類など)[4]。









- 甲殻亜門 Crustacea(側系統群)
- 貝形虫綱 Ostracoda - 貝虫(カイミジンコ、ウミホタルなど)
- ヒゲエビ亜綱[153] Mystacocarida - ヒゲエビ
- ウオヤドリエビ綱[153](イクチオストラカ綱[154]) Ichthyostraca
- 鰓尾亜綱 Branchiura - 鰓尾類(チョウ/ウオジラミなど)
- 舌形亜綱 Pentastomida - シタムシ
- 軟甲綱 Malacostraca - シャコ、カニ、エビ、オキアミ、ダンゴムシなど
- 六幼生綱[153](六齢ノープリウス綱[154]) Hexanauplia
- 鰓脚綱 Branchiopoda - アルテミア、ホウネンエビ、カブトエビ、カイエビ、ミジンコなど
- カシラエビ綱 Cephalocarida - カシラエビ
- ムカデエビ綱 Remipedia - ムカデエビ










-

-
 様々な昆虫
様々な昆虫


絶滅した分類群
以下は上記の現存群(鋏角類・多足類・甲殻類・六脚類)のクラウングループに含まれていない(例えば該当現存群に対して基盤的な化石分類群、ステムグループ)、もしくは所属する現存群が不確実な化石分類群を列挙する。
- ラディオドンタ目(放射歯目)Radiodonta - ラディオドンタ類[156][157][158][159](放射歯類[151]、広義のアノマロカリス類[160][161])。アノマロカリス、アンプレクトベルア、ペイトイア、フルディアなど。
-

-

-

-

-
 様々なラディオドンタ類
様々なラディオドンタ類





-

-
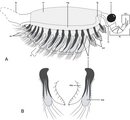 スルシカリス
スルシカリス


- (目)Hymenocarina[140]
- カナダスピス、ブランキオカリス、オダライア、ワプティアなど。真節足動物[163][164][22]/大顎類[140][142][165][110][166][167][162]/汎甲殻類[142][168][169]のいずれかのステムグループ。カンブリア紀に生息。
-

-

-
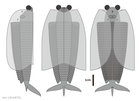
-
 ブランキオカリス
ブランキオカリス -
 トクンミア
トクンミア





- フーシェンフイア目 Fuxianhuiida[22]
- フーシェンフイア類。フーシェンフイア、チェンジャンゴカリスなど。真節足動物[163][164][20][22]/大顎類[140][142][110]のいずれかのステムグループ。カンブリア紀に生息。
-
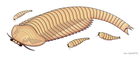
-

-
 アラカリス
アラカリス -
 グァンウェイカリス
グァンウェイカリス




- メガケイラ綱 Megacheira[145]
- メガケイラ類[156](大付属肢節足動物、大付属肢型節足動物)。ヨホイア、ハイコウカリス、レアンコイリア、フォルティフォルケプスなど。真節足動物[20][22][170][169][165]/鋏角類[141][110][162][66]のいずれかのステムグループ。カンブリア紀( - デボン紀?)に生息。





-
 クンミンゲラ
クンミンゲラ -
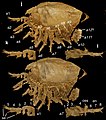 インディアナ(化石)
インディアナ(化石)


- (目)Phosphatocopina
- Hesslandona、Vestrogothia など。大顎類のステムグループ[164]もしくは甲殻類[172]。カンブリア紀に生息。

- マーレロモルフ綱 Marrellomorpha
- マーレロモルフ類[156][158](マレロモルフ類[159])。マーレラ、ヴァコシニア、フルカ、ミメタスターなど。真節足動物[140]/大顎類[164][170]のいずれかのステムグループ。カンブリア紀 - デボン紀に生息。
-

-
 ヴァコシニア
ヴァコシニア -
 フルカ
フルカ -
 ミメタスター
ミメタスター




- ユーシカルシノイド綱 Euthycarcinoidea
- ユーシカルシノイド類[156]。ユーシカルシヌス、ヘテロクラニア、アパンクラなど。大顎類[140]/多足類[173]のいずれかのステムグループ。カンブリア紀 - 三畳紀に生息。
-
 アパンクラ
アパンクラ -



- 嚢頭綱 Thylacocephala
- 嚢頭類(ティラコセファルス類[156])。Thylacares、ドロカリス、アンキトカゾカリス[174]など。甲殻類としての位置は不確実[175]。オルドビス紀[176](カンブリア紀?)- 白亜紀に生息。
-

-

-
 コンカヴィカリス
コンカヴィカリス -
 ドロカリス
ドロカリス




-

-
 Brittaniclus rankini
Brittaniclus rankini




- モリソニア目 Mollisoniida[179]
- モリソニア類。モリソニア[180]、セリオペなど。鋏角類/真鋏角類のいずれかのステムグループ[168][179]。カンブリア紀 - オルドビス紀に生息。
-

-
 セリオペ
セリオペ


- (亜門[181])Artiopoda[182][145]
- 三葉虫、光楯類など。真節足動物[140]/鋏角類[164][66]/大顎類[139][110][162]のいずれかのステムグループ。カンブリア紀 - ペルム紀に生息。
- (上綱[181])Vicissicaudata - シドネイア、エメラルデラ、ケロニエロン類、光楯類など。
- ケロニエロン目 Cheloniellida - ケロニエロン類。ケロニエロン[180]、ドゥスリアなど
- 光楯目 Aglaspidida - 光楯類(アグラスピス類)。アグラスピス、ベックウィジアなど。
- 三葉形類 Trilobitomorpha
- Petalopleura - シンダレラ、シャンダレラなど。
- Nektaspida - ナラオイアなど。
- Conciliterga - クアマイア、サペリオンなど。
- 三葉虫綱 Trilobita - 三葉虫。
-

-

-
 アグラスピス
アグラスピス -

-
 トリアルトルス(三葉虫)
トリアルトルス(三葉虫)





旧分類
分子系統学・分岐分類学が盛行する以前には、形態に基づく以下の分類体系が使用されていた。流通している書籍と文献にもこの分類にしたがっているものも多い。よって参考・比較のため、また生物学史上の意義もあり、以下に併記する。
- 大顎亜門 Mandibulata(その後の大顎類に相当)
節足動物と同じ種類の言葉
- 節足動物のページへのリンク