
ゴルゴノプス
| ゴルゴノプス | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 ゴルゴノプスの復元想像図
| ||||||||||||||||||||||||||||||
| 地質時代 | ||||||||||||||||||||||||||||||
| ペルム紀後期 | ||||||||||||||||||||||||||||||
| 分類 | ||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| 学名 | ||||||||||||||||||||||||||||||
| Gorgonops Owen, 1878 | ||||||||||||||||||||||||||||||
| 種 | ||||||||||||||||||||||||||||||
|
ゴルゴノプス (Gorgonops) は古生代ペルム紀後期の約2億4,800万年前に現在の南アフリカに生存していた単弓類の絶滅した属。属名の由来は、ギリシア神話のゴルゴーンから。1876年、リチャード・オーウェンによって命名された。模式種は G.torvus。当時としては極めて強力な捕食者であったが、ペルム紀末(P-T境界)の大絶滅を生き延びることはできなかった。
概要[編集]
ペルム紀後期タタール紀(またはロピンジアン期)のカルー盆地に繁栄した捕食者、ゴルゴノプス亜目の代表的な属である。全長は約2メートルで、ゴルゴノプス類において比較的大型の部類に入る。なお標準的なゴルゴノプス類は全長が1メートル前後とされる(参考はリカエノプス)[1]。
-
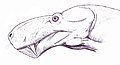 頭部復元図
頭部復元図 -
 頭部復元図
頭部復元図 -
 頭部骨格
頭部骨格



古生物学[編集]
頭部[編集]
下顎下端に達するほどの長い犬歯と発達した切歯をもち、顎関節は90度近く開いた[2]。これは長大な牙を相手に食い込ませるのに必要な進化だったとされる。犬歯の厚みは後の剣歯虎よりもやや厚いが、基本的にはサーベル状の薄く鋭利な構造だった[2]。ゴルゴノプスの歯は顎前方の切歯と犬歯が長く発達している代わりに、後方の頬歯(人間の奥歯にあたる歯)は短く退化気味である[1]。一部の近縁種では完全に頬歯が消失している。
ゴルゴノプスは、ディノケファルス類のようなそれまでの肉食獣弓類と異なり、下顎に明確な筋突起が発達し始めている[1]。これにより咬む力も強かったと思われる。とはいえ、より派生的な獣歯類のテロケファルス類やキノドン類よりも筋突起の発達具合は弱い。
脳の研究からゴルゴノプスを含むゴルゴノプス類は、通常姿勢では鼻先をやや下に傾斜させていた事が示されている[3]。
顎骨に多数の小さな窪みがあり、これは洞毛の痕跡ではないかといわれる。このことから[1][4]、既に体毛をもち、体温の保持ができたとも考えられる。当時のパンゲア大陸は半ば砂漠化しているような土地も多かったため、そのような地域での急激な気温の変化に対応するために体毛を獲得していた可能性がある。
四肢[編集]
ゴルゴノプスと彼らを含む初期の獣歯類は、それまでの獣弓類や爬虫類よりも脚全体が長くなっていた。一方で大腿骨は曲線を描いていたため、ギャロップ走行が可能だったとされている。こうしたギャロップ走法は必要に迫られた際にとった行動とされ、通常時はガニ股で爬行をしていた[5]。前足は依然として半直立だったが後ろ足は現代の哺乳類のように直立していたとみられる[6]。さらにゴルゴノプス類はそれまでの捕食者(ディノケファルス類)よりも頭部や牙を軽量化していたため、当時としてはかなり機動力に富む捕食者だったと考えられる。
胴体と内臓/代謝[編集]
ロバート・バッカーの恐竜異説によると、化石の産出状況(肉食の種類よりも植物食の種類が極端に豊富)や、形態的特徴(四肢や頭部の特徴)から、ゴルゴノプスを含む派生的な単弓類(獣弓類)は、それまでの基盤的な単弓類である盤竜類よりも代謝が高く、その代謝効率は現生哺乳類と盤竜類の中間に当たる。とされた。そして上記の考えにより、バッカー氏は獣弓類が気温の下がる冬においてでさえ、活動的に獲物を探していたとしている[7]。
ただし上記の研究は化石化のプロセス等を考慮しきれていないとして疑問符が投げかけられる事もある[1]。
古生態学[編集]
食性[編集]
ゴルゴノプスおよびゴルゴノプス類は生粋の肉食動物だった。糞化石の研究によると本種は、上記の身体的特徴を用いてディノケファルス類の生き残りと大小のディキノドン類のような数々の単弓類を狙い[8]、さらに側爬虫類のパレイアサウルス類や初期の双弓類のヨンギナといった爬虫類も襲った[8]。さらに水中の魚類や両生類(例は迷歯類)も漁っていた可能性がある[8]。
一つの可能性としては、同時代のパレイアサウルス類の多くが皮骨性の装甲(現生のワニの鎧に近い構造)をまとっていたため、長大な犬歯はそれらの外皮を貫いて仕留めるための進化だったのではないかと推測されている[1]。
ただし別の研究では、ゴルゴノプスおよびゴルゴノプス類は比較的小型の動物を丸呑みしていたのでは、とする報告も上がっている[9][8]。また近縁属のアエルログナトゥスの化石より、その腹部から小型のディキノドン類の化石が発見されている[10]。
一方で間接的な証拠である糞化石(生痕化石)より[8]、ゴルゴノプス類は小型の獲物は丸呑みにし、大型の獲物は骨を噛み砕いて飲み込んでいたともされている。この研究を受けサイエンスライターの金子降一氏は、自著において『ゴルゴノプス類の顎の力はハイエナ並みだった。』と推測している[1]。しかし現時点でゴルゴノプス類の咬筋の研究において、彼らの顎がハイエナに匹敵する力を秘めていたかどうかは不明である。
古環境[編集]
ゴルゴノプスの化石が発掘されたカルー盆地はペルム紀後期当時は、広大な河川堆積平野であった。ゴルゴノプスはじめ多くの化石(植物、昆虫、魚、更に爬虫類や単弓類といった有羊膜類)が、当時あったゴンドワニデ山脈(en:Gondwanide orogeny)から流れ出た巨大な河川によって堆積した地層(カルー超層群)から発見されている。当時、地球上の全ての大陸が合わさったことで超大陸パンゲア大陸が形成されていたのに加え、気温は温暖化しており、パンゲアの多くの地域では内陸部を中心に乾燥した大地が広がっていたが、ここでは豊かな水系に育まれたことで多くの動植物が繁栄していた。
絶滅[編集]
P/T境界(もしくは直前)の環境変化によりゴルゴノプスは絶滅した。本属の生態的地位(ニッチ)は、それ以降モスコリヌスのような肉食性テロケファルス類に引き継がれるも、ペルム紀後期に肉食性獣弓類の占有していた頂点捕食者のニッチには、三畳紀以降は肉食性主竜類が進出していくことになる。
分布[編集]
南アフリカのカルー盆地などから化石が産出している。2010年に発表された内容によれば、およそ90体もの化石が見つかっている[11]。
脚注[編集]
- ^ a b c d e f g 金子隆一『哺乳類型爬虫類―ヒトの知られざる祖先』朝日選書、1998年、204-214,246-247頁。ISBN 9784022597090。
- ^ a b 土屋健『カラー図解 古生物たちのふしぎな世界 繁栄と絶滅の古生代3億年史』講談社、2017年、222頁。ISBN 978-4-06-502018-0。
- ^ Ricardo Araújo; Vincent Fernandez; Michael J. Polcyn; Jörg Fröbisch; Rui M.S. Martins (2017-04-11). “Aspects of gorgonopsian paleobiology and evolution: insights from the basicranium, occiput, osseous labyrinth, vasculature, and neuroanatomy”. PeerJ. doi:10.7717/peerj.3119.
- ^ 成美堂出版 恐竜スーパーリアル大図鑑
- ^ Richard W. Blob (2001). “Evolution of hindlimb posture in nonmammalian therapsids: biomechanical tests of paleontological hypotheses”. Paleobiology 27 (1): 14-38. doi:10.1666/0094-8373(2001)027<0014:EOHPIN>2.0.CO;2.
- ^ 土屋健『石炭紀・ペルム紀の生物』技術評論社、2014年、112-113頁。ISBN 978-4-7741-6588-2。
- ^ ロバート・T・バッカー 著、瀬戸口烈司 訳『恐竜異説』平凡社、1989年。ISBN 4582524044。
- ^ a b c d e Roger M.H.Smith; Jennifer Botha-Brinkcd (2011). “Morphology and composition of bone-bearing coprolites from the Late Permian Beaufort Group, Karoo Basin, South Africa”. Palaeogeography, Palaeoclimatology, Palaeoecology 312 (1-2): 40-53. doi:10.1016/j.palaeo.2011.09.006.
- ^ Christian F. Kammerer (2016). “Systematics of the Rubidgeinae (Therapsida: Gorgonopsia)”. PeerJ. doi:10.7717/peerj.1608.
- ^ Michael W. Maisch (2009). “The small dicynodont Katumbia parringtoni (VON HUENE, 1942) (Therapsida:Dicynodontia) from the Upper Permian Kawinga Formation of Tanzania as gorgonopsian prey.”. Palaeodiversity 2: 279–282.
- ^ Merrill Nicolas; Bruce S. Rubidge (2010). “Changes in Permo-Triassic terrestrial tetrapod ecological representation in the Beaufort Group (Karoo Supergroup) of South Africa”. Lethaia 43 (1). doi:10.1111/j.1502-3931.2009.00171.x.
参考文献[編集]
書籍[編集]
- ティム・ヘインズ、ポール・チェンバーズ 著、椿正晴 訳『よみがえる恐竜・古生物』SBクリエイティブ、2006年7月15日、44-45頁。ISBN 4-7973-3547-5。
- グレゴリー・S・ポール 著、小畠郁生 訳『肉食恐竜事典』河出書房新社、1993年。ISBN 978-4309250618。(獣弓類やバッカーの研究に関連するページ)
- トーマス・R・ホルツ、トマス・V・レイ『ホルツ博士の最新恐竜事典』朝倉書店、2010年。ISBN 9784254162639。(バッカーの研究に関連するページ)
論文[編集]
- Blaire Van Valkenburgh; Ian Jenkins (2002). “Evolutionary Patterns in the History of Permo-Triassic and Cenozoic Synapsid Predators”. The Paleontological Society Papers 8.
- Eva V. I. Gebauer (2007). Phylogeny and Evolution of the Gorgonopsia with a Special Reference to the Skull and Skeleton of GPIT/RE/7113 ('Aelurognathus?' parringtoni).
関連項目[編集]
この項目は、古生物学に関連した書きかけの項目です。この項目を加筆・訂正などしてくださる協力者を求めています(プロジェクト:地球科学、プロジェクト:生物/Portal:地球科学、Portal:生物学)。 |