
Abstract
Rationale
Many animal models relevant to the persistent effects of drugs of abuse necessitate the application of interspecies dose scaling procedures to approximate drug administration regimens in humans, but drug self-administration procedures differ in that they allow animal subjects to control their own drug intake.
Objectives
This report reviews the reinforcing effects of 3,4-methylenedioxymethamphetamine (MDMA), its enantiomers, and several structural analogs in rhesus monkeys, paying particular attention to the pharmacological mechanisms of such reinforcing effects, the development of structure activity relationships among these compounds, the stability of MDMA self-administration behavior over time, and the persistent effects of self-administered MDMA on monoamines.
Results
The methylenedioxy amphetamine congeners MDMA, 3,4-methylenedioxyamphetamine, N-ethyl-3,4-methylenedioxyamphetamine, and N-methyl-1-(3,4-methylenedioxyphenyl)-2-butanamine function as reinforcers in rhesus monkeys, maintaining self-administration behavior greater than that engendered by contingent saline but less than that engendered by traditional psychostimulants. These findings are remarkable as structurally distinct serotonergic hallucinogen-like drugs do not maintain reliable self-administration in laboratory animals. During prolonged MDMA self-administration, MDMA-maintained responding progressively weakens, and MDMA eventually fails to maintain significant self-administration. The neurochemical correlates of this effect have not yet been identified.
Conclusions
Procedures in which MDMA and related compounds are self-administered can be established in rhesus monkeys. These techniques can be used to engender contingent MDMA exposure without resorting to controversial methods of interspecies dose scaling. As such, further application of self-administration methods may provide important new insights into the persistent effects of MDMA on brain and behavior in nonhuman primates.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The use of the term neurotoxicity to describe the persistent effects of the entactogen (Nichols 1986) 3,4-methylenedioxymethamphetamine (MDMA, “ecstasy”, Fig. 1a) has generally been criticized on two major grounds: the lack of a consistent definition for the term itself, and the questionable validity of interspecies dose scaling procedures used to equate experimental drug regimens in laboratory animals with recreational MDMA use in humans (see, e.g., Saunders 1995; Grob 2000; Kalia 2000; Vollenweider et al. 2001; Kish 2002; de la Torre and Farre 2004). With regard to the terminology dispute, there are at least three morphological criteria that are generally accepted as validated indicators of neural damage: frank cell loss, argyrophilia (silver staining), and reactive gliosis (O’Callaghan and Miller 1993). Current technological limitations dictate that all three of these measures be performed directly on brain tissue samples; thus, their application to human drug users has been sparse. Indeed, at the time of this writing, there exist no published reports describing frank cell loss, argyrophilia, or reactive gliosis in postmortem human brain tissue from MDMA users. As such, some researchers have relied instead on more ambiguous measures of axon terminal integrity in human MDMA users, such as depletion of serotonin (5-HT) content (see, e.g., Kish et al. 2000), reduced binding to plasmalemmal 5-HT transporters (see, e.g., McCann et al. 1998, 2005; Semple et al. 1999), or compensatory upregulation of postsynaptic 5-HT receptors (see, e.g., Reneman et al. 2002). Other researchers have eschewed the study of human drug users in favor of animal models, which frequently involved the application of various interspecies dose scaling procedures.

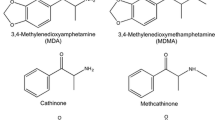
Chemical structures of the compounds studied in self-administration experiments. a 3,4-methylenedioxymethamphetamine (MDMA). b 3,4-methylenedioxyamphetamine (MDA). c N-ethyl-3,4-methylenedioxyamphetamine (MDE). d N-methyl-1-(3,4-methylenedioxyphenyl)-2-butanamine (MBDB)
Interspecies scaling is typically accomplished via one of two techniques. Mathematically complex physiological scaling methods applied to laboratory animals are useful in predicting many pharmacokinetic parameters in humans for a given drug but require costly and time consuming measurements of multiple variables, such as organ size, blood flow rates, tissue to blood partitioning, plasma protein binding, enzymatic kinetic parameters, and other metabolic factors such as drug absorption and clearance (Mahmood and Balian 1999; Parrott et al. 2005). The construction of such physiological models is thus unlikely to occur outside industrial settings, which largely precludes their development with relation to drugs of abuse. In contrast, allometric scaling approaches are less intricate, using a simple power function plotting body weight or surface area against some pharmacokinetic variable on a log–log scale. Such allometric models, however, can be influenced – sometimes profoundly so – by multiple factors, such as experimental design, species studied, analytical errors, interlaboratory variation, and the body weight range of the species used to construct the model (Mahmood and Balian 1999). Indeed, it has been reported that a tenfold range of estimates may be derived depending on which pharmacokinetic and corrective factors are thought relevant; thus, the validity of these models depends critically on the availability of sufficient data (Yates and Kugler 1986). The term for this mismatch between predicted and observed pharmacokinetics is “vertical allometry,” which has been demonstrated with many drugs (Mahmood and Balian 1999), some of which (for example, diazepam) have reinforcing effects in primate self-administration procedures (Bergman and Johanson 1985; Gomez et al. 2002). Since it is difficult to predict whether a drug will exhibit vertical allometry (Mahmood and Balian 1999), the reliance on pharmacokinetic parameters to scale from one species to another may be unsound. Finally, allometric scaling is often irrelevant when metabolism produces active metabolites or pharmacokinetics of other than first order (Mahmood and Balian 1999), which is particularly problematic with regard to MDMA as nonlinear pharmacokinetics have been reported in humans (de la Torre et al. 2000) and laboratory primates (Mechan et al. 2006), and at least 14 different metabolites are generated from this compound in vivo (Green et al. 2003).
The drug regimen most commonly used in laboratory primate studies of the persistent effects of MDMA involves “binge dosing,” or the repeated administration of two or more large doses per day, typically for four consecutive days. This dosing procedure was established in the primate by Ricaurte et al. (1985) with 3,4-methylenedioxyamphetamine (MDA, an analog and metabolite of MDMA, Fig. 1b), then extended to MDMA in subsequent studies (Ricaurte et al. 1988), and generally results in profound regional depletions in 5-HT, its metabolite 5-hydroxyindoleacetic acid, and decreased binding potential at serotonin transporters (SERT). Most other laboratory groups studying the effects of MDMA in primates also seem to have adopted this dosing procedure (see, for example, Slikker et al. 1988, 1989; Kleven et al. 1989; DeSouza et al. 1990; Ali et al. 1993; Colado et al. 1993; Frederick et al. 1998; O’Shea et al. 1998; Taffe et al. 2002; Bowyer et al. 2003). The consistency with which these serotonergic alterations are observed after repeated administrations of MDMA suggests that this compound certainly possesses the capacity to alter neurochemistry in a persistent manner, but considerable controversy exists surrounding the applicability of the dose regimen commonly utilized to induce these effects in comparison with recreational use of MDMA by humans. Among human users, during a given night of drug taking, a consumption of two to three 60–150 mg (Kirsch 1996; Wolf et al. 1995; Cole et al. 2002) ecstasy pills is typical. Some users may of course consume more pills, but less than 10% of human MDMA users are exposed to the drug more frequently than once per week (Winstock et al. 2001). Thus, on its face, the widely used “binge dosing” procedure in laboratory primates seems quite unlike the typical drug exposure pattern observed in humans. However, application of some allometric interspecies scaling procedures indicate that this particular drug regimen may be analogous to human MDMA use in recreational settings (Morgan 2000), although convincing arguments to the contrary have also been advanced (Vollenweider et al. 2001).
Recently, some researchers have proposed that effect scaling (see, e.g., Wang et al. 2004) may be a particularly useful method for equating experimental MDMA doses in laboratory animals with recreational MDMA doses in humans. The principle behind interspecies effect scaling is that drug doses which induce identical effects (behavioral, physiological, neurochemical, etc.) in two different species may be considered equivalent, and therefore directly compared across these particular species, even though these doses will likely differ on a milligram per kilogram basis. This paradigm may have particular resonance with behavioral pharmacologists, as it is similar in many ways to the common practice of comparing “behaviorally equivalent” doses across drugs. For example, doses of different drugs that engender identical self-administration behavior may be compared in further studies without the confound of rate dependence (see, for example, a comparison of the reinforcing effects of cocaine and R(−)-MDMA after antagonist challenge in Fantegrossi et al. 2002). Indeed, if the question at hand is whether MDMA is neurotoxic at the doses in which it is self-administered by humans, then a thorough evaluation of the effects of self-administered MDMA on various neurochemical and neuroanatomical endpoints in laboratory primates might be particularly relevant. However, before such studies can be undertaken, it is critical to first demonstrate that MDMA and its structural analogs can maintain self-administration behavior.
Reinforcing effects of MDMA and its analogs
Behaviorism proposes that the actions of an organism are governed by their consequences according to principles of operant conditioning (Skinner 1938; Ferster and Skinner 1957). To the extent that different routes of drug administration can influence onset of drug action, a majority of researchers using drug self-administration procedures have chosen to use intravenous preparations to maximize the speed with which drug effects are induced, although it should be noted that delivery of some compounds via intramuscular (see, e.g., Goldberg et al. 1976), inhalation (Carroll et al. 1990), and oral (Gomez et al. 2002) routes also maintains behavior in rhesus monkeys. In laboratory primates, intravenous preparations involve the surgical insertion of an indwelling venous catheter (typically into a femoral or jugular vein) that is then attached to a drug supply via an electronic infusion pump, the operation of which is controlled by the behavior of the animal according to experimenter-determined operant contingencies.
Previous research has amply demonstrated that response-dependent drug administration can have vastly different effects than one might predict based upon data gathered after noncontingent drug administration, and this principle seems to apply across drug classes. It is thus important to note that the reliance on experimenter-administered MDMA regimens in studies of persistent neurochemical effects in laboratory animals raises the question of whether these effects would be obtained via MDMA self-administration. For example, in the baboon, a history of midazolam self-administration resulted in potentiated sensitivity to the drug’s discriminative stimulus properties, while a history of response-independent midazolam decreased sensitivity to these same effects (Ator and Griffiths 1992). Among the psychostimulants, the lethal effects of cocaine were profoundly attenuated in rats self-administering the drug, as compared to a yoked control group receiving the same pattern of infusions noncontingently (Dworkin et al. 1995). Finally, there is evidence that the contingencies governing drug administration can even affect neurochemical measures. In this regard, Hemby et al. (1997) demonstrated significantly higher extracellular dopamine concentrations in the nucleus accumbens, despite equivalent brain concentrations of drug, in rats self-administering cocaine, as compared to a yoked control group. Furthermore, Stefanski et al. (1999) observed region-specific downregulations of D1 and D2 receptors in rat brain after a 5-week regimen of methamphetamine self-administration but not after equivalent response-independent drug exposure using a yoked control design. Based upon these and other data, the relevance of neurotoxicity studies relying on noncontingent MDMA administration to situations involving drug self-administration (such as that occurring in human ecstasy users) must be regarded as unknown.
Despite the widespread recreational use of MDMA, the reinforcing effects of ring-substituted amphetamine derivatives have not been extensively characterized in laboratory primates, although racemic MDMA has been shown to act as a reinforcer in intravenous self-administration paradigms in baboons (Lamb and Griffiths 1987) and rhesus monkeys (Beardsley et al. 1986; Fantegrossi et al. 2002; Lile et al. 2005). In addition to MDMA, the racemic forms of several structural analogs of MDMA, most notably N-ethyl-3,4-methylenedioxyamphetamine (MDE, Fig. 1c) (Sannerud et al. 1996) and MDA (Griffiths et al. 1976), have also been shown to maintain self-administration in baboons. This is an important point to consider, as the serotonergic hallucinogens of phenylisopropylamine and indolealkylamine structures have long been distinguished as drugs that are abused by humans but fail to engender reliable self-administration behavior in laboratory animals (Poling and Bryceland 1979; Fantegrossi et al. 2004b). It is interesting to note that hallucinogen-like compounds with antagonist affinity at glutamatergic N-methyl-d-aspartate receptors maintain self-administration behavior in rhesus monkeys, as reinforcing effects have been previously demonstrated with phencyclidine (Balster and Woolverton 1980), ketamine (Moreton et al. 1977), and memantine (Nicholson et al. 1998).
Although intruiging stereoselective effects have been described for the methylenedioxy amphetamine congeners in drug discrimination experiments (see, e.g., Glennon et al. 1982; Schechter 1987; Oberlender and Nichols 1988; Baker et al. 1995, 1997; Baker and Taylor 1997), the enantiomers have been studied in self-administration preparations far more rarely. A direct comparison of several of these compounds and their stereoisomers in the same group of animals, under the same self-administration contingencies, might reveal important structure–activity relationships (see, e.g., Anderson et al. 1978; Nichols 1981) for the reinforcing effects of MDMA-like drugs. Such a study has not yet been conducted, but we have studied a small sample of these compounds in rhesus monkey self-administration paradigms, and a review of these data may be illustrative.
MDMA self-administration
Procedure
The reinforcing effects of racemic MDMA and its enantiomers were compared to those of cocaine and methamphetamine in studies previously reported (Fantegrossi et al. 2002). In these studies, as well as those reported below, monkeys had extensive pharmacological histories including self-administration of various psychostimulants and opioids for several years before initiation of studies involving the reinforcing effects of the entactogens. Self-administration behavior was assessed in two 60-min experimental sessions conducted each day: a morning session starting at 10:00 h and an afternoon session starting at 16:00 h. The onset of each session was signaled by illumination of a red stimulus light. In the presence of this light, the tenth response on the lever beneath it resulted in the operation of the infusion pump (FR10). During the 5-s infusion duration, the red stimulus light was extinguished, the center green light was illuminated, and further lever presses had no programmed consequences. Immediately after the termination of each infusion, all stimulus lights were extinguished for a 1-min time out period (TO 1 min) during which lever presses continued to have no programmed consequences. Each TO period counted toward the total 60-min session time. Under baseline conditions, animals were maintained on a cocaine dose of 0.01 mg/kg per injection following the above outlined behavioral requirements. To ensure that responding was being maintained by the presented drug, saline was randomly substituted for cocaine approximately every third or fourth session, usually for two consecutive sessions. MDMA substitutions occurred approximately twice per week, and no substitutions were made on weekends. All drug substitutions followed an ascending dose order within drug, and at least four recovery sessions occurred between substitution trials.
Results and discussion
In these studies, all the three forms of MDMA dose-dependently maintained self-administration behavior that was less than that engendered by cocaine or methamphetamine but greater than that engendered by contingent saline infusions (Fantegrossi et al. 2002). These results are similar to those obtained in a recent progressive ratio study (Lile et al. 2005) where MDMA maintained significantly lower break points than did cocaine. Although R(−)-MDMA exhibits markedly lower activity in an assay of inhibition of dopamine uptake (Steele et al. 1987), and a 25-fold lower affinity for the dopamine transporter than its S(+)-enantiomer (Setola et al. 2003), no obvious differences in drug potency or effectiveness in maintaining contingent responding have been observed for the MDMA stereoisomers in self-administration experiments. Indeed, the finding that R(−)-MDMA functions as a reinforcer at all is puzzling.
5-HT antagonists and MDMA self-administration
Procedure
Further studies with the 5-HT2A/2C antagonist ketanserin and the selective 5-HT2A antagonist M100907 (formerly MDL100907) were also conducted to probe the pharmacological mechanisms underlying the reinforcing effects of the MDMA enantiomers (Fantegrossi et al. 2002). Monkeys were tested twice daily on a 130-min FR30 TO 45 s session maintained by four discrete doses of either IV cocaine, S(+)-MDMA, or R(−)-MDMA. This schedule is quite similar to that previously described by Winger et al. (1989, 1992). Each component of this multiple schedule was followed by a 10-min blackout period during which all stimulus lights were turned off and responses had no programmed consequences. The onset of each component was signaled by the illumination of the red stimulus light. In the presence of this light, the 30th response on the lever beneath it (FR 30) resulted in operation of the infusion pump. During the infusion, the red stimulus light was extinguished and the center green light was illuminated. Each component of this multiple schedule allowed a maximum of 20 self-injections, or the lapse of 25 min. Infusions were followed by a 45-s inter-injection timeout period (TO 45 s) during which all lights were turned off and responses had no programmed consequences.
Discrete dose components differed from each other in the duration of infusion pump operation. The pumps were calibrated to deliver 1 ml of solution over a 5-s interval; thus, by lengthening the infusion interval, a greater amount of solution was administered, thereby providing a higher dose per infusion. Throughout these experiments, the pump durations were 0.5, 1.7, 5.0, and 16.7 s. These durations corresponded to doses of 0.001, 0.003, 0.01, and 0.03 mg/kg per injection when drug reservoir supply bags were filled with a concentration of 0.01 mg kg−1 ml−1 (supply bags were made on an individual basis, dependent on the animal’s weight.) Two higher doses were tested by filling supply bags with a concentration of 0.1 mg kg−1 ml−1 (again made on an individual basis, dependent on the animal’s weight.) Under these conditions, the pump durations remained the same but corresponded to doses of 0.01, 0.03, 0.1, and 0.3 mg/kg per injection. The overlapping doses of 0.01 and 0.03 were thus delivered at two different infusion durations, allowing a comparison of the effect of infusion duration on response rates. Four different orders of pump durations were used: an ascending order, a descending order, and two mixed orders. One of these orders was selected randomly before each cocaine or saline session; however, all results are based upon data obtained under the ascending order schedule. Saline was substituted for the baseline drug (cocaine) approximately every third session. During saline substitution sessions, saline was available during all four components, and rates of responding were required to be ≤0.5 response per second, or else saline was delivered contingently on subsequent sessions until this response requirement was met. The MDMA stereoisomers were substituted for cocaine periodically, but no more often than every fourth session. MDMA substitutions and 5-HT2A antagonist pretreatments were always conducted under the ascending dose order. Pretreatments with the 5-HT2A antagonists ketanserin and M100907 were administered intramuscularly (IM) 15-min before the start of the session.
Results and discussion
These antagonist studies suggested that the reinforcing effects of MDMA were serotonergically mediated (Fantegrossi et al. 2002). This effect was particularly pronounced for R(−)-MDMA, as responding for this enantiomer was completely abolished by M100907 at a dose that did not alter cocaine-maintained behavior. It is interesting to note that the reinforcing effects of S(+)-MDMA were somewhat more resistant to disruption by these antagonists.
Stability of MDMA self-administration
Procedure
Subsequent self-administration experiments with MDMA were conducted over an approximately 18-month period to assess the persistence of these reinforcing effects (Fantegrossi et al. 2004a) because diminished subjective effects over repeated uses have often been reported by human ecstasy users (see, for example, the recent review of chronic tolerance to MDMA in Parrot 2005). In these studies, monkeys were given the opportunity to self-administer one of several MDMA doses about twice per week under the single-dose FR10 schedule described above, and dose–effect curves were repeatedly determined in this manner over the duration of the experiment.
Results and discussion
Comparing the first and last dose–effect curves for racemic and R(−)-MDMA generally revealed shifts to the right and down, indicating reduced reinforcing effects for these compounds (Fantegrossi et al. 2004a). For S(+)-MDMA, however, decreased reinforcing effects were observed in only one monkey (Fantegrossi et al. 2004a). It is important that responding for cocaine was remarkably stable in all monkeys, suggesting that there was some pharmacological specificity to these observed behavioral changes.
It has been suggested that these data may indicate the development of chronic tolerance to the effects of MDMA (Parrot 2005); however, it may be more difficult to assert that tolerance has developed to the reinforcing effects of a drug than is commonly appreciated. In the case of complete tolerance to the reinforcing effects of a drug, it seems reasonable to assume that the drug would lose its capacity to engender self-administration (as no amount of drug could be taken that could serve the function of maintaining the behavior) and extinction would thus ensue. On the other hand, if some incomplete tolerance were to occur, then one might predict that drug taking would actually increase to compensate for the decreased sensitivity to the reinforcing effects of that drug. In both instances, the problem with interpreting changes in self-administration behavior in terms of tolerance is to determine whether such tolerance truly applies to the reinforcing effects of the drug or to some other concomitant drug effect. The previously described progressive loss of MDMA self-administration can thus be taken to indicate either complete tolerance to the reinforcing effects of the drug or complete loss of reinforcing effect for some other unknown reason. It is clear that further investigation into the persistence of MDMA-maintained behavior is warranted.
MDA self-administration
Procedure
Racemic MDA and its enantiomers were studied in five monkeys with extensive histories of intravenous drug self-administration, using the multiple-dose FR30 schedule described above. In brief, this schedule presented opportunities to self-administer up to 20 injections in each of the four discrete dose components (where dose is changed by altering infusion duration). The primary dependent variable under this schedule is response rate, and because total number of injections per dose component is capped at 20, the ongoing rate of behavior is the preferred of index of reinforcing effect.
Results and discussion
Mean rates of responding for contingent saline were low at all infusion durations. They are indicated by the shaded region in Fig. 2. In contrast, racemic MDA (Fig. 2, filled diamonds) and S(+)-MDA (Fig. 2, filled triangles) engendered high rates of dose-dependent responding, generating “inverted U”-shaped dose–effect functions similar to those commonly observed with other drug reinforcers. Overall drug intake for these compounds was high, and subjects earned a majority of available injections across the dose range tested. It is interesting to note that on average, R(−)-MDA (Fig. 2, open inverted triangles) maintained low rates across the dose range tested and did not engender responding significantly different from contingent saline at any dose. Drug intake was generally quite low for this enantiomer, and few available infusions were earned in each dose component. However, as was previously observed with some indolealkylamine and phenylisopropylamine hallucinogens (Fantegrossi et al. 2004b), there were discrete substitution sessions where high rates of response were engendered in individual monkeys. Response rates far in excess of one response per second were intermittently observed in two monkeys when the high dose range of R(−)-MDA was available, and in these particular sessions, almost all available infusions were earned (Fig. 3). This pattern of responding is consistent with the notion of “transient reinforcing effects”, which we have previously suggested to be characteristic of the more traditional serotonergic hallucinogens (see Fantegrossi et al. 2004b for an extended discussion). These findings suggest similarities between R(−)-MDA and the classical psychedelics beyond those previously demonstrated in terms of discriminative stimulus effects (see, e.g., Glennon et al. 1982; Baker and Taylor 1997).

Self-administration dose–effect curves for S(+)-MDA (open triangles), racemic MDA (filled diamonds), and R(−)-MDA (inverted open triangles), and the range of response rates for contingent saline (shaded region) under a multiple-dose FR30 schedule. Each point represents the mean (n=5) response rate±SEM. Abscissa: dose of self-administered drug (milligram per kilogram per injection). Ordinate: rate, in responses per second. See text for further details

Transient reinforcing effects of R(−)-MDA in monkeys 96X4394 (left panel) and 98X3152 (right panel) during high dose range substitutions. Each bar represents the absolute response rate for a single dose within a single substitution session; numerals above each bar indicate the number of infusions earned (maximum = 20) for that dose component. Abscissae: dose of self-administered drug (milligram per kilogram per injection). Ordinates: rate, in responses per second. See text for further details
MDE self-administration
Procedure
MDE is the short-acting (Shulgin and Shulgin 1991) N-ethyl homolog of MDMA. This compound occasions MDMA-appropriate responding (Boja and Schechter 1987; Glennon and Misenheimer 1989; Schechter 1998) but does not share discriminative stimulus effects with the phenylisopropylamine hallucinogen 1-(2,5-dimethoxy-4-methylphenyl)-2-aminopropane (DOM) or amphetamine (Glennon et al. 1988). MDE is frequently present in illicit pills sold as “ecstasy” (Cole et al. 2002). Racemic MDE and its enantiomers were studied in four monkeys with extensive histories of intravenous drug self-administration, using the single-dose FR10 schedule described above. In brief, this schedule presented opportunities to repeatedly self-administer a single-unit dose of drug over a 60-min session. Under this procedure, the total number of injections earned is not limited but is governed solely by the subject’s behavior, and is thus the preferred index of reinforcing effects.
Results and discussion
The mean number of contingent saline infusions earned under this schedule was low. It is indicated by the shaded region in Fig. 4. Racemic MDE (Fig. 4, filled diamonds), S(+)-MDE (Fig. 4, filled triangles), and R(−)-MDE (Fig. 4, open inverted triangles) each dose-dependently engendered significant self-administration behavior under this schedule. It is interesting to note that S(+)-MDE was approximately tenfold more potent than the R(−) enantiomer, while the effective doses for the racemate were intermediate to those of the stereoisomers. These results are of the sort that one might intuitively expect to obtain when comparing the effects of a racemic mixture with those of its component isomers, but the fact that this pattern of data is obtained only rarely with regard to the reinforcing effects of the methylenedioxy amphetamine congeners is perhaps notable. In fact, the “disconcerting absence of additivity between the component isomers in comparison with the activity of the racemate” was demonstrated with MDMA in one of the earliest publications regarding the in vivo effects of these compounds (Anderson et al. 1978). Why MDE and its enantiomers seem to deviate from this pattern in terms of their reinforcing effects remains unknown.

Self-administration dose–effect curves for S(+)-MDE (open triangles), racemic MDE (filled diamonds), and R(−)-MDE (inverted open triangles), and the range of injections of contingent saline (shaded region) earned under a single-dose FR10 schedule. Each point represents the mean (n=3) number of injections earned±SEM. Abscissa: dose of self-administered drug (milligram per kilogram per injection). Ordinate: number of injections per session. See text for further details
MBDB self-administration
Procedure
N-methyl-1-(3,4-methylenedioxyphenyl)-2-butanamine (MBDB, Fig. 1d) is the alpha-ethyl homolog of MDMA. This compound is similar to MDMA in terms of inhibition of 5-HT uptake and stimulation of 5-HT release but has greatly reduced dopaminergic effects (Nichols 1986; Johnson et al. 1986). Furthermore, MBDB generalizes to MDMA but does not occasion LSD- or DOM-appropriate responding in drug discrimination tests (Oberlender and Nichols 1988). Although MBDB has previously been shown to induce conditioned place preference in the rat (Marona-Lewicka et al. 1996), the reinforcing effects of this compound have not previously been studied in any species using intravenous self-administration procedures. It was therefore of interest to compare the reinforcing effects of racemic MBDB to those of racemic MDMA in four monkeys with extensive histories of intravenous drug self-administration, using the single-dose FR10 schedule previously described in the MDMA and MDE sections above.
Results and discussion
As seen in the MDE experiments, the mean number of contingent saline infusions earned under this schedule was low. It is indicated by the shaded region in Fig. 5. Racemic MDMA (Fig. 5, filled diamonds) and racemic MBDB (Fig. 5, open circles) each dose-dependently engendered significant self-administration behavior under this schedule; however, MDMA was more potent and effective in maintaining high response rates than was MBDB. These results thus recapitulate the previous place preference experiments in which the conditioned effects of MBDB were at least 2.5-fold weaker than those of MDMA (Marona-Lewicka et al. 1996). Despite the reduced conditioned and reinforcing effects of the alpha-ethyl homolog as compared to the parent compound, moderate MBDB self-administration and conditioned place preference are nevertheless remarkable considering the almost complete lack of dopaminergic effects induced by this compound (Nichols 1986; Johnson et al. 1986, 1991; Nash and Nichols 1991; Marona-Lewicka et al. 1996). As seems to be the case with MDMA, particularly the R(−) enantiomer (Fantegrossi et al. 2002), the reinforcing effects of MBDB may be mediated through serotonergic mechanisms. Further study of MBDB (and other 5-HT-selective analogs of MDMA) using self-administration techniques may uncover novel pharmacological mediators of the reinforcing effects of the methylenedioxy amphetamine congeners.

Self-administration dose–effect curves for racemic MDMA (filled diamonds) and racemic MBDB (open circles), and the range of injections of contingent saline (shaded region) earned under a single-dose FR10 schedule. Each point represents the mean (n=2) number of injections earned±SEM. Abscissa: dose of self-administered drug (milligram per kilogram per injection). Ordinate: number of injections per session. See text for further details
Reinforcing effects—unresolved issues
The data described in the preceding sections and those published in previous reports demonstrate that MDMA and related entactogens maintain significant self-administration behavior in laboratory primates, although the various measures used to indicate reinforcing effects are generally lesser in magnitude with these compounds than with traditional psychostimulants such as cocaine and methamphetamine (Beardsley et al. 1986; Lamb and Griffiths 1987; Fantegrossi et al. 2002; Lile et al. 2005). Nevertheless, the fact that these drugs function as reinforcers offers researchers a procedure with which to study the persistent effects of these compounds on brain, behavior, and perhaps cognition. Furthermore, the seemingly temporary period during which MDMA maintains behavior may provide an absolute maximum drug intake which can be compared to the drug regimens administered noncontingently in standard neurotoxicity studies (see below). The pharmacological relevance of such experimenter-administered drug regimens must be called into question if they result in total MDMA doses comparable to or greater than the estimated “lifetime” intakes obtained in a self-administration setting, particularly if such regimens are completed in a matter of days.
The use of self-administration procedures in effect scaling, however, is more complicated than has been described so far. The techniques outlined in the studies above were not initially designed to assess long-term changes in drug intake, and the finding that the reinforcing effects of MDMA did not persist was neither anticipated nor initially welcomed. However, when it became clear that MDMA-maintained behavior was indeed decreasing in magnitude, every attempt was made to characterize this phenomenon with some degree of rigor and precision. Although this single demonstration of the effect has yet to be systematically replicated, important questions are raised nonetheless.
First, how specific is this behavioral change across the entactogens, and can the persistence (or lack thereof) of reinforcing effects for these compounds be related to pharmacological activity? The finding that the reinforcing effects of S(+)-MDMA were more resistant to disruption than were those of R(−)-MDMA suggests that some structure–activity relationships may emerge with more thorough study of the phenomenon and highlights the importance of studying the enantiomers in this context. Similarly, what operant contingencies impact the rate of degradation of the reinforcing effects of MDMA? In this regard, it seems reasonable to assume that more frequent access to higher unit doses of drug would hasten the loss of reinforcing effects, while more intermittent exposure to lower doses may prolong the capacity of MDMA to reinforce behavior; however, at this point, one can only speculate.
As “schedule-dependence” is a robust phenomenon in behavioral pharmacology (for reviews, see, e.g., Woods et al. 1975; Sanger and Blackman 1976; LeSage et al. 1999), the particulars of the operant conditioning schedules used to engender contingent responding for the entactogens are likely to be critical mediators of the manifestation of this effect. Depending on the particulars of one’s experimental questions, self-administration paradigms involving limited (e.g., see Roberts and Andrews 1997; Tornatzky and Miczek 1999) or continuous (see, e.g., Bozarth and Wise 1985; Tornatzky and Miczek 2000) access to MDMA may be most useful. Finally, the behavioral and pharmacological histories of the subjects used in long-term MDMA self-administration studies are also likely to be relevant factors. In contrast to rodents, laboratory primates are rarely experimentally naïve, which is often viewed as a weakness in experimental design. However, it should be noted that human ecstasy users are notorious for their polydrug abuse (Pedersen and Skrondal 1999), and the current body of research describing the persistent effects of MDMA in humans has previously been criticized for its general lack of inclusion of polydrug abuser control subjects (Schifano et al. 1998; Kish 2002). Thus, the use of rhesus monkeys with extensive drug self-administration histories may actually have more validity in comparison to the human ecstasy user population than the use of drug-naïve subjects. However, the role of historical variables in the acquisition and long-term maintenance of MDMA self-administration certainly deserves thorough investigation.
Effects of self-administered MDMA on monoamines
While it is becoming increasingly common to couple dynamic in vivo extracellular fluid sampling techniques (such as voltammetry or microdialysis) with self-administration behavior to measure the acute effects of drugs on neurochemistry, the persistent effects of self-administered drugs on monoamines have not been intensively studied. Although a single report suggests that even a relatively brief period of MDA self-administration results in the depletion of hippocampal 5-HT at least 2 weeks after discontinuing drug access in the rat (Markert and Roberts 1991), the persistence of this effect was not determined, and only one previous study has examined the neurochemical consequences of self-administered MDMA (Fantegrossi et al. 2004a). The behavioral data obtained from this later study were presented above, but a significant effort was made to uncover neurochemical correlates of these observed behavioral changes, the results of which are described below.
The effects of experimenter-administered MDMA on monoamines have been reviewed previously (most recently in Itzhak and Achat-Mendes 2004; Baumgarten and Lachenmayer 2004; Reneman 2003; Green et al. 2003; Gudelsky and Yamamoto 2003; Lyles and Cadet 2003) and generally describe a profile consisting of extensive and persistent depletions of central 5-HT, reduced SERT expression, and inhibition of 5-HT synthesis in the absence of effects on other monoamines. Effects consistent with gliosis, however, may or may not be discussed. Nevertheless, as described above, behavioral pharmacology has demonstrated quite convincingly that the effects of drugs with diverse mechanisms of action can be profoundly influenced by the contingency (or noncontingency) of their administration. As such, the potential pitfalls of interspecies dose scaling notwithstanding, data obtained from animal neurotoxicity studies relying on noncontingent MDMA administration may not necessarily be congruent with results of experiments where subjects self-administer the drug.
In the long-term MDMA self-administration experiments described above, the selective reduction in the reinforcing effects of MDMA was not accompanied by significant depletion of 5-HT, dopamine, or their metabolites despite opportunities to self-administer MDMA and its enantiomers on at least 120 separate occasions. In comparison, the aforementioned drug regimen commonly invoked to induce enduring neural changes in nonhuman primates results in a daily exposure to 10 to 20 mg/kg MDMA for four consecutive days. In contrast, self-administered MDMA intakes were between approximately 120 and 250 mg/kg (the sum of all self-administered intakes of racemic MDMA or its enantiomers) over the 18-month duration of the experiment. Thus, the presently discussed lack of monoamine depletions in the MDMA self-administration animals cannot simply be due to insufficient MDMA intake. Nevertheless, acute MDMA self-administration sessions for each rhesus monkey occurred where drug intakes of 6–15 mg/kg were observed; these doses approach or exceed the amounts typically administered noncontingently to primates in other studies in a single day. If the effects of noncontingent MDMA exposure are cumulative, as seems to be assumed by the broadly used 4-day dosing regimen, one may presume that the effects of contingent MDMA administration would be cumulative as well. On the other hand, the present results might argue that a key toxic level of MDMA (or a metabolite) must be achieved with each administration to induce persistent neural effects. Thus, with regard to frank neurotoxicity, total amount of MDMA administered may be a less important factor than duration and pattern of MDMA exposure, although a definitive statement in this regard would require more empirical data than have thus far been collected. Finally, it should be noted that all monkeys in the presently discussed self-administration studies – even those in the MDMA-naïve control group – had extensive drug self-administration histories, which may have resulted in neurochemical changes that could mask any effect of self-administered MDMA. In addition, there are surprisingly few data on baseline monoamine levels in adult rhesus monkeys, so it is difficult to know how much variability to expect between subjects.
In this regard, we studied tissue samples from additional 19 monkeys with histories of drug self-administration and noted a fairly wide range of serotonin and dopamine concentrations in MDMA-naïve subjects (see filled circles in Fig. 6). The monoamine concentrations of four monkeys with long-term MDMA self-administration histories (Fig. 6, open circles) fall within this range, but whether these control animals accurately reflect basal monoamine levels in drug-naïve monkeys is unknown. At the time of this writing, we simply cannot say with certainty how short-term exposure to high doses of noncontingent MDMA (as in the standard nonhuman primate neurotoxicity studies) differs in a neurochemical sense, from long-term self-administration of relatively low doses of the drug. A prospective study comparing monoamine levels before and after initiating an MDMA self-administration regimen would be particularly informative in this regard.

Tissue concentrations of 5-HT (top) or dopamine (bottom) in caudate and putamen. Filled circles represent individual monoamine concentrations for MDMA-naïve control animals, open circles represent individual monoamine concentrations for monkeys with MDMA self-administration histories. Horizontal lines denote the mean values for each group±SEM. Abscissae: brain region. Ordinates: monoamine concentration (nanogram per milligram tissue). Brain concentrations of 5-HT and dopamine were measured by reverse phase HPLC coupled with electrochemical detection using the method previously described in Fantegrossi et al. (2004a). See text for further details
It seems reasonable to assume, however, that the persistent effects of MDMA would be related to the degree of occupancy achieved at the drug’s sites of action, the most obvious of which is the SERT (Rudnick and Wall 1992). It is of interest that in vivo neuroimaging of dopamine transporter (DAT) occupancy using positron emission tomography (PET) has revealed identical potencies (on a milligram per kilogram basis) for this measure with both intravenous cocaine and methylphenidate in three distinct species: mice, baboons, and humans (Gatley et al. 1999). These results suggest that interspecies dose scaling would not be necessary with these psychostimulants if one were interested in studying effects of these compounds mediated by DAT occupancy. It is presently unknown whether results of this nature would be recapitulated at the SERT with MDMA and its analogs, but this is an empirical question which could be addressed with a PET study of SERT occupancy using an appropriate radioligand.
Summary and conclusions
The use of self-administration procedures to study the behavioral, neurochemical, and perhaps even neurotoxic effects of MDMA seems to have several advantages over noncontingent administration of the drug. First, allowing animals to regulate their own drug administration (within the necessary constraints imposed by the experimenter-determined operant schedule) may avoid the necessity of interspecies dose scaling and can result in intakes that seem to be more consistent with estimates of human MDMA consumption. Second, drug administration in a self-administration setting is much more similar to the conditions in which humans use MDMA, and although not yet explicitly demonstrated with the methylenedioxy amphetamine congeners, profound differences in drug effects as a function of the contingency or noncontingency of their delivery have been reported with other drug classes. Third, as described in this review, self-administration of MDMA and its analogs is a reasonably robust phenomenon, particularly in the rhesus monkey (but see De La Garza’s review in this issue for a discussion of the challenges in maintaining rodent self-administration of MDMA). Finally, the progressive loss of MDMA self-administration behavior previously noted in rhesus monkeys replicates the similar phenomenon of spontaneous abstinence in human users (von Sydow et al. 2002; Parrot 2005), and may be a novel and sensitive behavioral model for the persistent neurochemical effects of the drug. In approximately 20 years of MDMA research, only a handful of studies have employed self-administration techniques to study the effects of this compound. Increased application of these methods, particularly when coupled with neurochemical measurements or neuroimaging, might provide important new insights into the persistent effects of MDMA on brain and behavior.
References
Ali SF, Newport GD, Scallet AC, Binienda Z, Ferguson SA, Bailey JR, Paule MG, Slikker W Jr (1993) Oral administration of 3,4-methylenedioxymethamphetamine (MDMA) produces selective serotonergic depletion in the nonhuman primate. Neurotoxicol Teratol 15(2):91–96
Anderson GM 3rd, Braun G, Braun U, Nichols DE, Shulgin AT (1978) Absolute configuration and psychotomimetic activity. NIDA Res Monogr 22:8–15
Ator NA, Griffiths RR (1992) Differential sensitivity to midazolam discriminative-stimulus effects following self-administered versus response-independent midazolam. Psychopharmacology 110:1–4
Baker L, Taylor M (1997) Assessment of the MDA and MDMA optical isomers in a stimulant-hallucinogen discrimination. Pharmacol Biochem Behav 57(4):737–748
Baker LE, Broadbent J, Michael EK, Matthews PK, Metosh CA, Saunders RB, West WB, Appel JB (1995) Assessment of the discriminative stimulus effects of the optical isomers of ecstasy (3,4-methylenedioxymethamphetamine; MDMA). Behav Pharmacol 6(3):263–275
Baker LE, Virden TB, Miller ME, Sullivan CL (1997) Time course analysis of the discriminative stimulus effects of the optical isomers of 3,4-methylenedioxymethamphetamine (MDMA). Pharmacol Biochem Behav 58(2):505–516
Balster RL, Woolverton WL (1980) Continuous-access phencyclidine self-administration by rhesus monkeys leading to physical dependence. Psychopharmacology 70(1):5–10
Baumgarten HG, Lachenmayer L (2004) Serotonin neurotoxins—past and present. Neurotox Res 6(7–8):589–614
Beardsley PM, Balster RL, Harris LS (1986) Self-administration of methylenedioxymethamphetamine (MDMA) by rhesus monkeys. Drug Alcohol Depend 18(2):149–157
Bergman J, Johanson CE (1985) The reinforcing properties of diazepam under several conditions in the rhesus monkey. Psychopharmacology 86(1–2):108–113
Boja JW, Schechter MD (1987) Behavioral effects of N-ethyl-3,4-methylenedioxyamphetamine (MDE; “EVE”). Pharmacol Biochem Behav 28(2):153–156
Bowyer JF, Young JF, Slikker W, Itzak Y, Mayorga AJ, Newport GD, Ali SF, Frederick DL, Paule MG (2003) Plasma levels of parent compound and metabolites after doses of either d-fenfluramine or d-3,4-methylenedioxymethamphetamine (MDMA) that produce long-term serotonergic alterations. Neurotoxicology 24:379–390
Bozarth MA, Wise RA (1985) Toxicity associated with long-term intravenous heroin and cocaine self-administration in the rat. JAMA 254(1):81–83
Carroll ME, Krattiger KL, Gieske D, Sadoff DA (1990) Cocaine-base smoking in rhesus monkeys: reinforcing and physiological effects. Psychopharmacology 102:443–450
Colado MI, Murray TK, Green AR (1993) 5-HT loss in rat brain following 3,4-methylenedioxymethamphetamine (MDMA), p-chloroamphetamine, and fenfluramine administration and effects of chlormethiazole and dizocilpine. Br J Pharmacol 108:583–589
Cole JC, Bailey M, Sumnall HR, Wagstaff GF, King LA (2002) The content of ecstasy tablets: implications for the study of their long-term effects. Addiction 97(12):1531–1536
de la Torre R, Farre M (2004) Neurotoxicity of MDMA (ecstasy): the limitations of scaling from animals to humans. Trends Pharmacol Sci 25(10):505–508
de la Torre R, Farre M, Ortuno J, Mas M, Brenneisen R, Roset PN, Segura J, Cami J (2000) Non-linear pharmacokinetics of MDMA (‘ecstasy’) in humans. Br J Clin Pharmacol 49(2):104–109
DeSouza EB, Battaglia G, Insel TR (1990) Neurotoxic effects of MDMA on brain serotonin neurons: evidence from neurochemical and radioligand binding studies. Ann NY Acad Sci 600:682–698
Dworkin SI, Mirkis S, Smith JE (1995) Response-dependent versus response-independent presentation of cocaine: differences in the lethal effects of the drug. Psychopharmacology 117:262–266
Fantegrossi W, Ullrich T, Rice K, Woods J, Winger G (2002) 3,4-Methylenedioxymethamphetamine (MDMA,‘Ecstasy’) and its stereoisomers as reinforcers in rhesus monkeys: serotonergic involvement. Psychopharmacology 161:356–364
Fantegrossi WE, Woolverton WL, Kilbourn M, Sherman P, Yuan J, Hatzidimitriou G, Ricaurte GA, Woods JH, Winger G (2004a) Behavioral and neurochemical consequences of long-term intravenous self-administration of MDMA and its enantiomers by rhesus monkeys. Neuropsychopharmacology 29(7):1270–1281
Fantegrossi WE, Woods JH, Winger G (2004b) Transient reinforcing effects of phenylisopropylamine and indolealkylamine hallucinogens in rhesus monkeys. Behav Pharmacol 15(2):149–157
Ferster CB, Skinner BF (1957) Schedules of reinforcement. Appleton, New York
Frederick DL, Ali SF, Gillam MP, Gossett J, Slikker W, Paule MG (1998) Acute effects of dexfenfluramine (d-FEN) and methylenedioxymethamphetamine (MDMA) before and after short-course, high-dose treatment. Ann NY Acad Sci 844:183–190
Gatley SJ, Volkow ND, Gifford AN, Fowler JS, Dewey SL, Ding YS, Logan J (1999) Dopamine-transporter occupancy after intravenous doses of cocaine and methylphenidate in mice and humans. Psychopharmacology 146:93–100
Glennon RA, Misenheimer BR (1989) Stimulus effects of N-monoethyl-1-(3,4-methylenedioxyphenyl)-2-aminopropane (MDE) and N-hydroxy-1-(3,4-methylenedioxyphenyl)-2-aminopropane (N-OH MDA) in rats trained to discriminate MDMA from saline. Pharmacol Biochem Behav 33(4):909–912
Glennon RA, Young R, Rosecrans JA, Anderson GM (1982) Discriminative stimulus properties of MDA analogs. Biol Psychiatry 17:807–814
Glennon RA, Yousif M, Patrick G (1988) Stimulus properties of 1-(3,4-methylenedioxyphenyl)-2-aminopropane (MDA) analogs. Pharmacol Biochem Behav 29(3):443–449
Goldberg SR, Morse WH, Goldberg DM (1976) Behavior maintained under a second-order schedule by intramuscular injection of morphine or cocaine in rhesus monkeys. J Pharmacol Exp Ther 199:278–286
Gomez TH, Roache JD, Meisch RA (2002) Orally delivered alprazolam, diazepam, and triazolam as reinforcers in rhesus monkeys. Psychopharmacology 161(1):86–94
Green AR, Mechan AO, Elliott JM, O’Shea E, Colado MI (2003) The pharmacology and clinical pharmacology of 3,4-methylenedioxymethamphetamine (MDMA, “ecstasy”). Pharmacol Rev 55(3):463–508
Griffiths RR, Winger G, Brady JV, Snell JD (1976) Comparison of behavior maintained by infusions of eight phenylethylamines in baboons. Psychopharmacology 50(3):251–258
Grob CS (2000) Deconstructing ecstasy: the politics of MDMA research. Addict Res 8:549–588
Gudelsky GA, Yamamoto BK (2003) Neuropharmacology and neurotoxicity of 3,4-methylenedioxymethamphetamine. Methods Mol Med 79:55–73
Hemby SE, Co C, Koves TR, Smith JE, Dworkin SI (1997) Differences in extracellular dopamine concentrations in the nucleus accumbens during response-dependent and response-independent cocaine administration in the rat. Psychopharmacology 133:7–16
Itzhak Y, Achat-Mendes C (2004) Methamphetamine and MDMA (ecstasy) neurotoxicity: ‘of mice and men’. IUBMB Life 56(5):249–255
Johnson MP, Hoffman AJ, Nichols DE (1986) Effects of the enantiomers of MDA, MDMA and related analogues on [3H]serotonin and [3H]dopamine release from superfused rat brain slices. Eur J Pharmacol 132(2–3):269–276
Johnson MP, Conarty PF, Nichols DE (1991) [3H]monoamine releasing and uptake inhibition properties of 3,4-methylenedioxymethamphetamine and p-chloroamphetamine analogues. Eur J Pharmacol 200(1):9–16
Kalia M (2000) Do validated biological measures of neurotoxicity really support the claim that MDMA is neurotoxic to man? Neuropsychobiology 42:45
Kirsch MM (1996) Ecstasy. In: Designer drugs, CompCare, Minneapolis (Minn), pp 75–97
Kish SJ (2002) How strong is the evidence that brain serotonin neurons are damaged in human users of ecstasy? Pharmacol Biochem Beh 71:845–855
Kish SJ, Furukawa Y, Ang L, Vorce SP, Kalasinsky KS (2000) Striatal serotonin is depleted in brain of a human MDMA (Ecstasy) user. Neurology 55:294–296
Kleven MS, Woolverton WL, Seiden LS (1989) Evidence that both intragastric and subcutaneous administration of methylenedioxymethylamphetamine (MDMA) produce serotonin neurotoxicity in rhesus monkeys. Brain Res 488:121–125
Lamb RJ, Griffiths RR (1987) Self-injection of d,1-3,4-methylenedioxymethamphetamine (MDMA) in the baboon. Psychopharmacology 91(3):268–272
LeSage MG, Stafford D, Glowa JR (1999) Preclinical research on cocaine self-administration: environmental determinants and their interaction with pharmacological treatment. Neurosci Biobehav Rev 23(5):717–741
Lile JA, Ross JT, Nader MA (2005) A comparison of the reinforcing efficacy of 3,4-methylenedioxymethamphetamine (MDMA, “ecstasy”) with cocaine in rhesus monkeys. Drug Alcohol Depend 78(2):135–140
Lyles J, Cadet JL (2003) Methylenedioxymethamphetamine (MDMA, Ecstasy) neurotoxicity: cellular and molecular mechanisms. Brain Res Brain Res Rev 42(2):155–168
Mahmood I, Balian JD (1999) The pharmacokinetic principles behind scaling from preclinical results to phase I protocols. Clin Pharmacokinet 36(1):1–11
Markert LE, Roberts DC (1991) 3,4-Methylenedioxyamphetamine (MDA) self-administration and neurotoxicity. Pharmacol Biochem Behav 39(3):569–574
Marona-Lewicka D, Rhee GS, Sprague JE, Nichols DE (1996) Reinforcing effects of certain serotonin-releasing amphetamine derivatives. Pharmacol Biochem Behav 53(1):99–105
McCann UD, Szabo Z, Scheffel U, Dannals RF, Ricaurte GA (1998) Positron emission tomographic evidence of toxic effect of MDMA (“ecstasy”) on brain serotonin neurons in human beings. Lancet 352:1433–1437
McCann UD, Szabo Z, Seckin E, Rosenblatt P, Mathews WB, Ravert HT, Dannals RF, Ricaurte GA (2005) Quantitative PET studies of the serotonin transporter in MDMA users and controls using [11C]McN5652 and [11C]DASB. Neuropsychopharmacology 30(9):1741–1750. doi:10.1038/sj.npp.1300736
Mechan A, Yuan J, Hatzidimitriou G, Irvine RJ, McCann UD, Ricaurte GA (2006) Pharmacokinetic profile of single and repeated oral doses of MDMA in squirrel monkeys: relationship to lasting effects on brain serotonin neurons. Neuropsychopharmacology 31(2):339–350. doi:10.1038/sj.npp.1300808
Moreton JE, Meisch RA, Stark L, Thompson T (1977) Ketamine self-administration by the rhesus monkey. J Pharmacol Exp Ther 203(2):303–309
Morgan MJ (2000) Ecstasy (MDMA): a review of its possible persistent psychological effects. Psychopharmacology 152:230–248
Nash JF, Nichols DE (1991) Microdialysis studies on 3,4-methylenedioxyamphetamine and structurally related analogues. Eur J Pharmacol 200(1):53–58
Nichols DE (1981) Structure–activity relationships of phenethylamine hallucinogens. J Pharm Sci 70(8):839–849
Nichols DE (1986) Differences between the mechanism of action of MDMA, MBDB, and the classic hallucinogens. Identification of a new therapeutic class: entactogens. J Psychoactive Drugs 18(4):305–313
Nicholson KL, Jones HE, Balster RL (1998) Evaluation of the reinforcing and discriminative stimulus properties of the low-affinity N-methyl-d-aspartate channel blocker memantine. Behav Pharmacol 9(3):231–243
O’Callaghan JP, Miller DB (1993) Quantification of reactive gliosis as an approach to neurotoxicity assessment. NIDA Res Monogr 136:188–212
O’Shea E, Granados R, Esteban B, Colado MI, Green AR (1998) The relationship between the degree of neurodegeneration of rat brain 5-HT nerve terminals and the dose and frequency of administration of MDMA (‘Ecstasy’). Neuropharmacology 37:919–926
Oberlender R, Nichols DE (1988) Drug discrimination studies with MDMA and amphetamine. Psychopharmacology 95(1):71–76
Parrot AC (2005) Chronic tolerance to recreational MDMA (3,4-methylenedioxymethamphetamine) or ecstasy. J Psychopharmacol 19:75–87
Parrott N, Jones H, Paquereau N, Lave T (2005) Application of full physiological models for pharmaceutical drug candidate selection and extrapolation of pharmacokinetics to man. Basic Clin Pharmacol Toxicol 96:193–199
Pedersen W, Skrondal A (1999) Ecstasy and new patterns of drug use: a normal population study. Addiction 94(11):1695–1706
Poling A, Bryceland J (1979) Voluntary drug self-administration by nonhumans: a review. J Psychedelic Drugs 11:185–190
Reneman L (2003) Designer drugs: how dangerous are they? J Neural Transm Suppl (66):61–83
Reneman L, Endert E, de Bruin K, Lavalaye J, Feenstra MG, de Wolff FA, Booij J (2002) The acute and chronic effects of MDMA (“ecstasy”) on cortical 5-HT2A receptors in rat and human brain. Neuropsychopharmacology 26(3):387–396
Ricaurte GA, Bryan G, Strauss L, Seiden L, Schuster C (1985) Hallucinogenic amphetamine selectively destroys brain serotonin nerve terminals. Science 229:986–988
Ricaurte GA, DeLanney LE, Irwin I, Langston JW (1988) Toxic effects of MDMA on central serotonergic neurons in the primate: importance of route and frequency of drug administration. Brain Res 446(1):165–168
Roberts DC, Andrews MM (1997) Baclofen suppression of cocaine self-administration: demonstration using a discrete trials procedure. Psychopharmacology 131(3):271–277
Rudnick G, Wall S (1992) The molecular mechanism of [3,4-methylenedioxy-methamphetamine (MDMA)]: serotonin transporters are targets for MDMA-induced serotonin release. Proc Natl Acad Sci U S A 89:1817–1821
Sanger DJ, Blackman DE (1976) Rate-dependent effects of drugs: a review of the literature. Pharmacol Biochem Behav 4(1):73–83
Sannerud CA, Kaminski BJ, Griffiths RR (1996) Intravenous self-injection of four novel phenethylamines in baboons. Behav Pharmacol 7(4):315–323
Saunders N (1995) Ecstasy and the dance culture. Neal’s Yard Desktop, London
Schechter MD (1987) MDMA as a discriminative stimulus: isomeric comparisons. Pharmacol Biochem Behav 27(1):41–44
Schechter MD (1998) MDMA-like stimulus effects of hallucinogens in male Fawn-Hooded rats. Pharmacol Biochem Behav 59(2):265–270
Schifano F, Di Furia L, Forza G, Minicuci N, Bricolo R (1998) MDMA (‘ecstasy’) consumption in the context of polydrug abuse: a report on 150 patients. Drug Alcohol Depend 52(1):85–90
Semple DM, Ebmeier KP, Glabus MF, O’Carroll RE, Johnstone EC (1999) Reduced in vivo binding to the serotonin transporter in the cerebral cortex of MDMA (“ecstasy”) users. Br J Psychiatry 175:63–69
Setola V, Hufeisen SJ, Grande-Allen KJ, Vesely I, Glennon RA, Blough B, Rothman RB, Roth BL (2003) 3,4-methylenedioxymethamphetamine (MDMA, “Ecstasy”) induces fenfluramine-like proliferative actions on human cardiac valvular interstitial cells in vitro. Mol Pharmacol 63(6):1223–1229
Shulgin A, Shulgin A (1991) PIHKAL: a chemical love story. Transform, Berkley, pp. 727–739
Skinner BF (1938) The behavior of organisms. Appleton, New York
Slikker W Jr, Ali SF, Scallet AC, Frith CH, Newport GD, Bailey JR (1988) Neurochemical and neurohistological alterations in the rat and monkey produced by orally administered methylenedioxymethamphetamine (MDMA). Toxicol Appl Pharmacol 94(3):448–457
Slikker W Jr, Holson RR, Ali SF, Kolta MG, Paule MG, Scallet AC, McMillan DE, Bailey JR, Hong JS, Scalzo FM (1989) Behavioral and neurochemical effects of orally administered MDMA in the rodent and nonhuman primate. Neurotoxicology 10:529–542
Steele TD, Nichols DE, Yim GK (1987) Stereochemical effects of 3,4-methylenedioxymethamphetamine (MDMA) and related amphetamine derivatives on inhibition of uptake of [3H]monoamines into synaptosomes from different regions of rat brain. Biochem Pharmacol 36(14):2297–2303
Stefanski R, Ladenheim B, Lee SH, Cadet JL, Goldberg SR (1999) Neuroadaptations in the dopaminergic system after active self-administration but not after passive administration of methamphetamine. Eur J Pharmacol 371:123–135
Taffe MA, Davis SA, Yuan J, Schroeder R, Hatzidimitriou G, Parsons LH, Ricaurte GA, Gold LH (2002) Cognitive performance of MDMA-treated rhesus monkeys: sensitivity to serotonergic challenge. Neuropsychopharmacology 27(6):993–1005
Tornatzky W, Miczek KA (1999) Repeated limited access to i.v. cocaine self-administration: conditioned autonomic rhythmicity illustrating “predictive homeostasis”. Psychopharmacology 145(2):144–152
Tornatzky W, Miczek KA (2000) Cocaine self-administration “binges”: transition from behavioral and autonomic regulation toward homeostatic dysregulation in rats. Psychopharmacology 148(3):289–298
Vollenweider FX, Jones RT, Baggott MJ (2001) Caveat emptor: editors beware (reply). Neuropsychopharmacology 24(4):461–463
von Sydow K, Lieb R, Pfister H, Hofler M and Wittchen HU (2002) Use, abuse and dependence of ecstacy and related drugs in adolescents and young adults—a transit phenomenon? Results from a longitudinal community study. Drug Alcohol Depend 66(2):147–159
Wang X, Baumann MH, Xu H, Rothman RB (2004) 3,4-Methylenedioxymethamphetamine (MDMA) administration to rats decreases brain tissue serotonin but not serotonin transporter protein and glial fibrillary acidic protein. Synapse 53:240–248
Winger G, Palmer R, Woods J (1989) Drug-reinforced responding: rapid determination of dose-response functions. Drug Alcohol Depend 24:135–142
Winger G, Skjoldager P, Woods J (1992) Effects of buprenorphine and other opioid agonists and antagonists on alfentanil- and cocaine-reinforced responding in rhesus monkeys. J Pharmacol Exp Ther 261:311–317
Winstock AR, Griffiths P, Stewart D (2001) Drugs and the dance music scene: a survey of current drug use patterns among a sample of dance music enthusiasts in the UK. Drug Alcohol Depend 64(1):9–17
Wolf K, Hay AWM, Sherlock K, Conner M (1995) Contents of ‘ecstasy’. Lancet 346:1100–1101
Woods JH, Downs DA, Carney J (1975) Behavioral functions of narcotic antagonists: response-drug contingencies. Fed Proc 34(9):1777–1784
Yates FE, Kugler PN (1986) Similarity principles and intrinsic geometries: contrasting approaches to interspecies scaling. J Pharm Sci 75(11):1019–1027
Acknowledgements
Expert technical assistance with monkey self-administration experiments was provided by Erich Doser, Debbie Huntzinger, Sarah Pilkington, and Jolan Terner at the University of Michigan. The author expresses his gratitude to Mike Kilbourne, Phil Sherman, Kyle Kuzspit, and Leslie Doherty at the University of Michigan PET facility for their skilled aid in the conduct of PET imaging procedures. This topic was presented at the 2005 annual meeting of the International Study Group Investigating Drugs as Reinforcers, and the insightful comments of those in attendance were incorporated into the resulting manuscript. An early draft of this review was read with perspicacity by Leonard Howell and Gail Winger, and the author thanks them for their editorial guidance. The studies described in this report were supported by USPHS Grants DA09161, DA05923, DA05707, and DA00206.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Fantegrossi, W.E. Reinforcing effects of methylenedioxy amphetamine congeners in rhesus monkeys: are intravenous self-administration experiments relevant to MDMA neurotoxicity?. Psychopharmacology 189, 471–482 (2007). https://doi.org/10.1007/s00213-006-0320-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-006-0320-8